

Balancing selection in the wild: testing population genetics theory of self-incompatibility in the rare species Brassica insularis
- PMID: 15944365
- PMCID: PMC1456519
- DOI: 10.1534/genetics.104.035915
Balancing selection in the wild: testing population genetics theory of self-incompatibility in the rare species Brassica insularis
Abstract
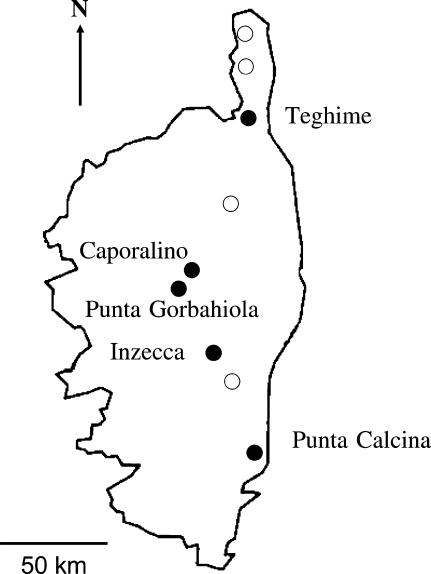
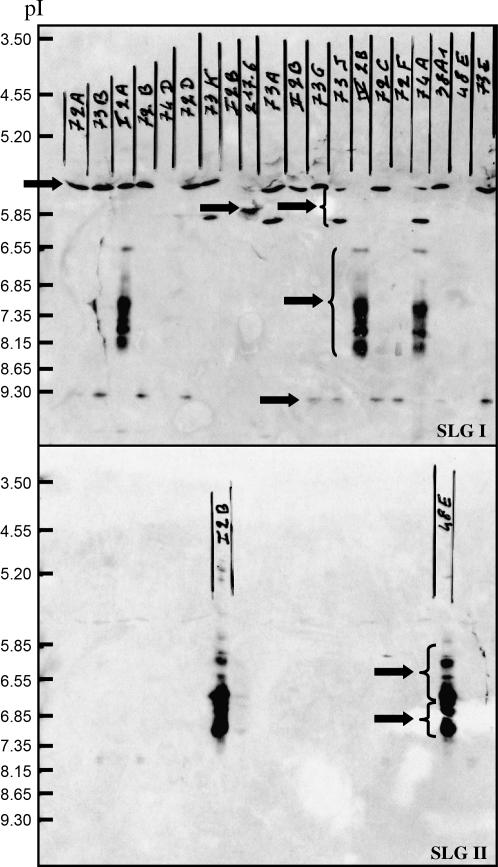
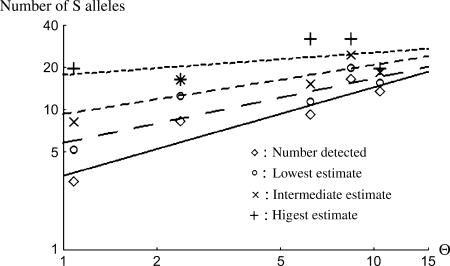
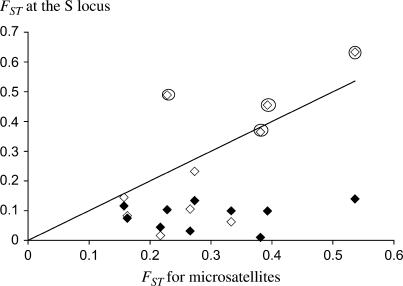
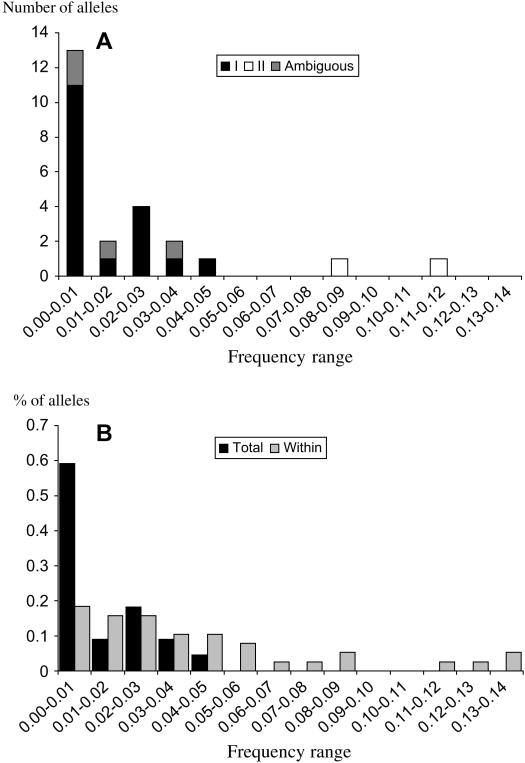
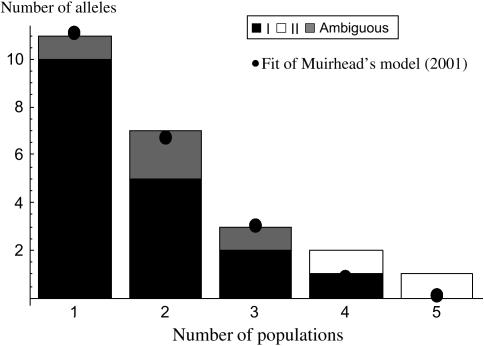
Self-incompatibility (SI) systems are widespread mechanisms that prevent self-fertilization in angiosperms. They are generally encoded by one genome region containing several multiallelic genes, usually called the S-locus. They involve a recognition step between the pollen and the pistil component and pollen is rejected when it shares alleles with the pistil. The direct consequence is that rare alleles are favored, such that the S-alleles are subject to negative frequency-dependent selection. Several theoretical articles have predicted the specific patterns of polymorphism, compared to neutral loci, expected for such genes under balancing selection. For instance, many more alleles should be maintained and populations should be less differentiated than for neutral loci. However, empirical tests of these predictions in natural populations have remained scarce. Here, we compare the genetic structure at the S-locus and microsatellite markers for five natural populations of the rare species Brassica insularis. As in other Brassica species, B. insularis has a sporophytic SI system for which molecular markers are available. Our results match well the theoretical predictions and constitute the first general comparison of S-allele and neutral polymorphism.
Figures

References
-
- Aspinwall, N., and T. Christian, 1992. Pollination biology, seed production, and population structure in queen-of-the-prairie, Filipendula rubra (Rosaceae) at Botkin fen, Missouri. Am. J. Bot. 79: 488–494.
-
- Bechsgaard, J., T. Bataillon and M. H. Schierup, 2004. Uneven segregation of sporophytic self-incompatibility alleles in Arabidopsis lyrata. J. Evol. Biol. 17: 554–561. - PubMed
-
- Bell, C. J., and J. R. Ecker, 1994. Assignment of 30 microsatellite loci to the linkage map of Arabidopsis. Genomics 19: 137–144. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
