

Evolutionary diversity and potential recombinogenic role of integration targets of Non-LTR retrotransposons
- PMID: 15944437
- PMCID: PMC1400617
- DOI: 10.1093/molbev/msi188
Evolutionary diversity and potential recombinogenic role of integration targets of Non-LTR retrotransposons
Abstract
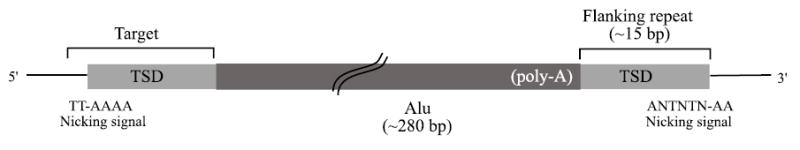
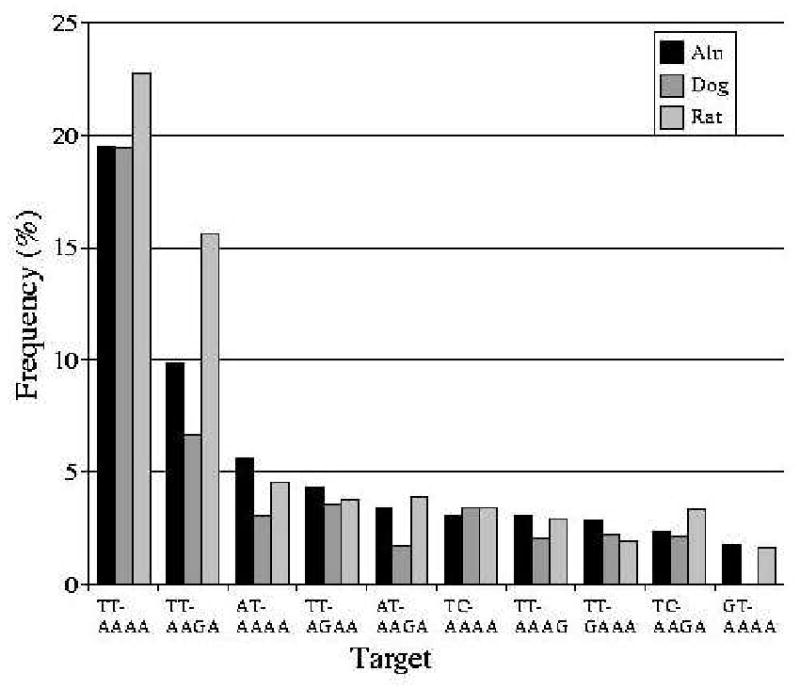
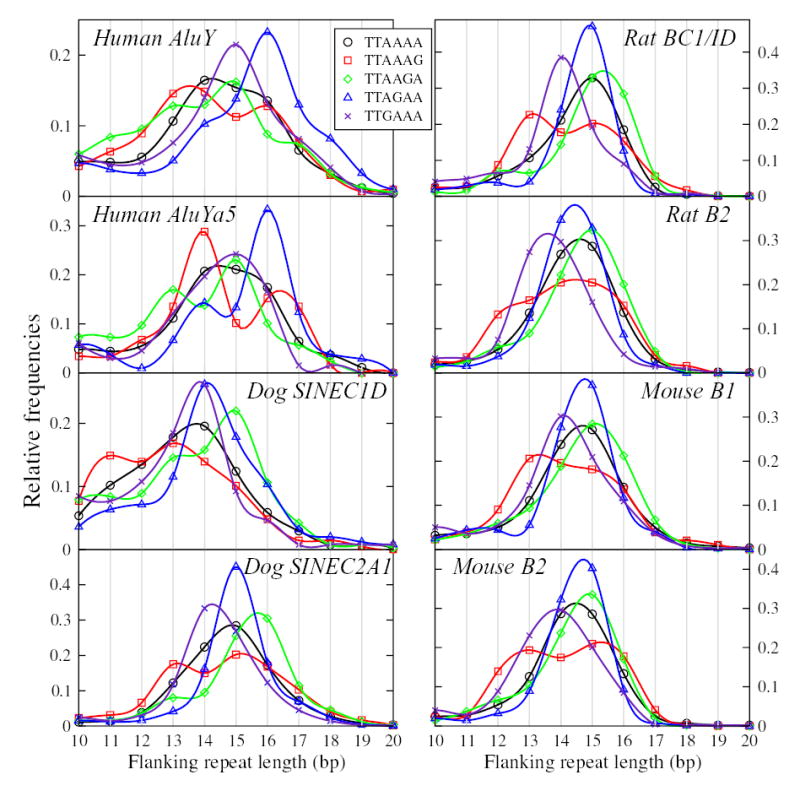
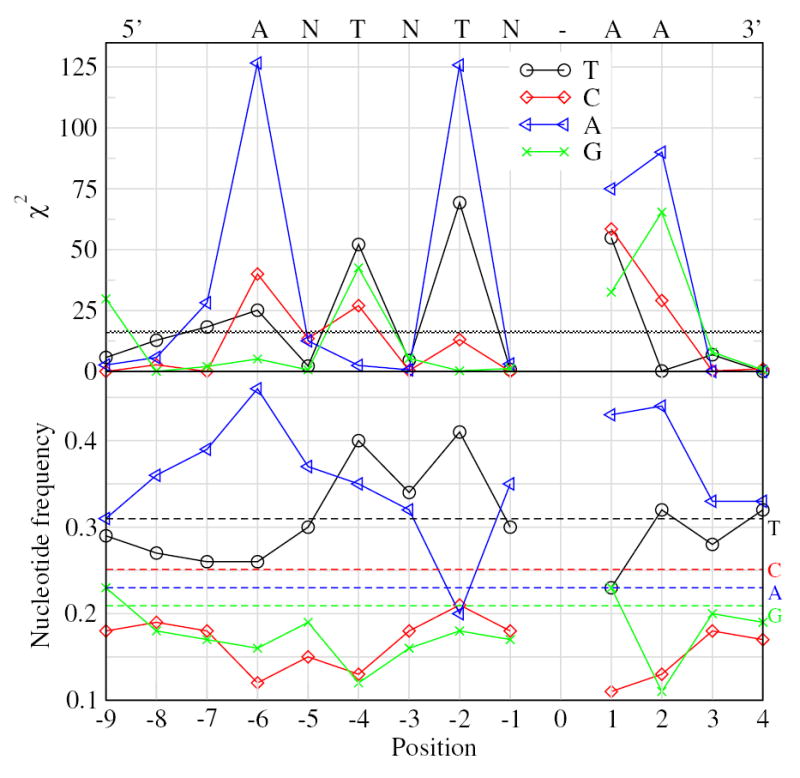
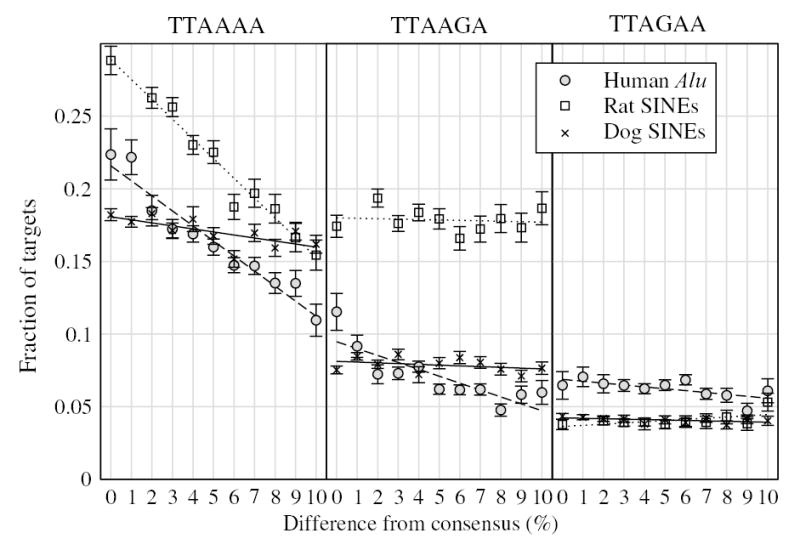
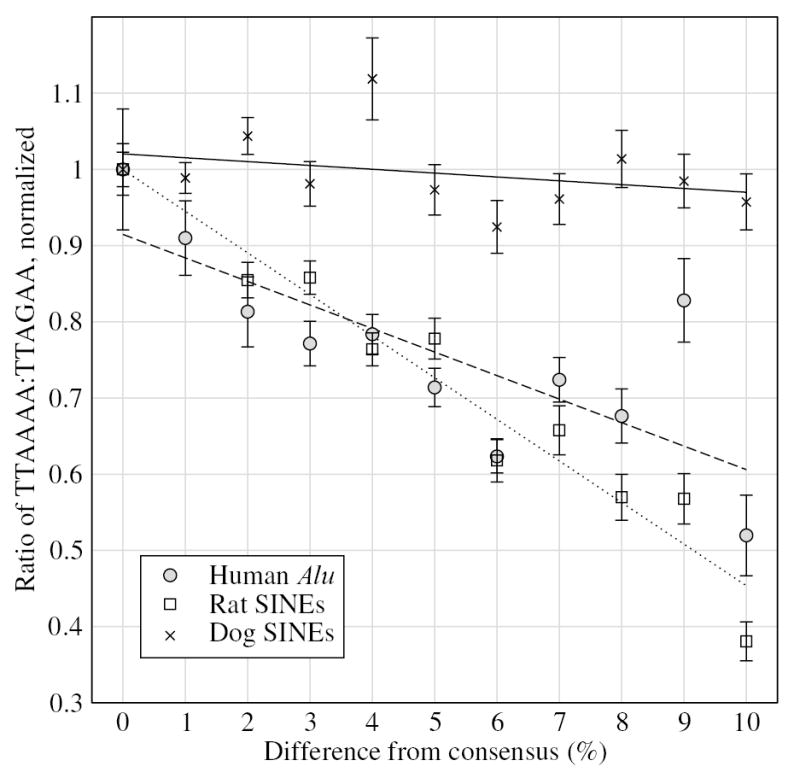
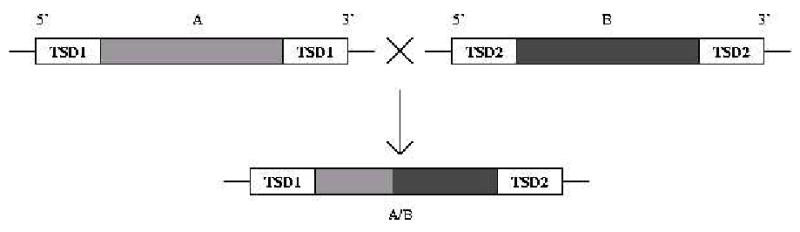
Short interspersed elements (SINEs) make up a significant fraction of total DNA in mammalian genomes, providing a rich substrate for chromosomal rearrangements by SINE-SINE recombinations. Proliferation of mammalian SINEs is mediated primarily by long interspersed element 1 (L1) non-long terminal repeat retrotransposons that preferentially integrate at DNA sequence targets with an average length of approximately 15 bp and containing conserved endonucleolytic nicking signals at both ends. We report that sequence variations in the first of the two nicking signals, represented by a 5'-TT-AAAA consensus sequence, affect the position of the second signal thus leading to target site duplications (TSDs) of different lengths. The length distribution of TSDs appears to be affected also by L1-encoded enzyme variants because targets with the same 5' nicking site can be of different average lengths in different mammalian species. Taking this into account, we reanalyzed the second nicking site and found that it is larger and includes more conserved sites than previously appreciated, with a consensus of 5'-ANTNTN-AA. We also studied potential involvement of the nicking sites in stimulating recombinations between SINEs. We determined that SINEs retaining TSDs with perfect 5'-TT-AAAA nicking sites appear to be lost relatively rapidly from the human and rat genomes and less rapidly from dog. We speculate that the introduction of DNA breaks induced by recurring endonucleolytic attacks at these sites, combined with the ubiquitousness of SINEs, may significantly promote recombination between repetitive elements, leading to the observed losses. At the same time, new L1 subfamilies may be selected for "incompatibility" with preexisting targets. This provides a possible driving force for the continual emergence of new L1 subfamilies which, in turn, may affect selection of L1-dependent SINE subfamilies.
Figures
Similar articles
-
Targeted identification of short interspersed nuclear element families shows their widespread existence and extreme heterogeneity in plant genomes.Plant Cell. 2011 Sep;23(9):3117-28. doi: 10.1105/tpc.111.088682. Epub 2011 Sep 9. Plant Cell. 2011. PMID: 21908723 Free PMC article.
-
Novel SINE families from salmons validate Parahucho (Salmonidae) as a distinct genus and give evidence that SINEs can incorporate LINE-related 3'-tails of other SINEs.Mol Biol Evol. 2007 Aug;24(8):1656-66. doi: 10.1093/molbev/msm083. Epub 2007 Apr 29. Mol Biol Evol. 2007. PMID: 17470437
-
[Non-LTR retrotransposons: LINEs and SINEs in plant genome].Yi Chuan. 2006 Jun;28(6):731-6. Yi Chuan. 2006. PMID: 16818439 Review. Chinese.
-
The conserved 3' Angio-domain defines a superfamily of short interspersed nuclear elements (SINEs) in higher plants.Plant J. 2020 Feb;101(3):681-699. doi: 10.1111/tpj.14567. Epub 2019 Dec 3. Plant J. 2020. PMID: 31610059
-
Retroposons of salmonoid fishes (Actinopterygii: Salmonoidei) and their evolution.Gene. 2009 Apr 1;434(1-2):16-28. doi: 10.1016/j.gene.2008.04.022. Epub 2008 May 22. Gene. 2009. PMID: 18590946 Review.
Cited by
-
The human LINE-1 retrotransposon creates DNA double-strand breaks.J Mol Biol. 2006 Apr 14;357(5):1383-93. doi: 10.1016/j.jmb.2006.01.089. Epub 2006 Feb 9. J Mol Biol. 2006. PMID: 16490214 Free PMC article.
-
Carnivore-specific SINEs (Can-SINEs): distribution, evolution, and genomic impact.J Hered. 2011 Sep-Oct;102 Suppl 1(Suppl 1):S2-10. doi: 10.1093/jhered/esr051. J Hered. 2011. PMID: 21846743 Free PMC article. Review.
-
The dynamic proliferation of CanSINEs mirrors the complex evolution of Feliforms.BMC Evol Biol. 2014 Jun 20;14:137. doi: 10.1186/1471-2148-14-137. BMC Evol Biol. 2014. PMID: 24947429 Free PMC article.
-
Inviting instability: Transposable elements, double-strand breaks, and the maintenance of genome integrity.Mutat Res. 2007 Mar 1;616(1-2):46-59. doi: 10.1016/j.mrfmmm.2006.11.021. Epub 2006 Dec 8. Mutat Res. 2007. PMID: 17157332 Free PMC article. Review.
-
Characterization of pre-insertion loci of de novo L1 insertions.Gene. 2007 Apr 1;390(1-2):190-8. doi: 10.1016/j.gene.2006.08.024. Epub 2006 Sep 12. Gene. 2007. PMID: 17067767 Free PMC article.
References
-
- Bentolila S, Bach JM, Kessler JL, Bordelais I, Cruaud C, Weissenbach J, Panthier JJ. Analysis of major repetitive DNA sequences in the dog (Canis familiaris) genome. Mamm Genome. 1999;10:699–705. - PubMed
-
- Brosius, J. 2005. Echoes from the past - are we still in an RNP world ? Cytogenet. Genome Res., In press. - PubMed
