

Human Rad51 filaments on double- and single-stranded DNA: correlating regular and irregular forms with recombination function
- PMID: 15944450
- PMCID: PMC1145190
- DOI: 10.1093/nar/gki640
Human Rad51 filaments on double- and single-stranded DNA: correlating regular and irregular forms with recombination function
Abstract
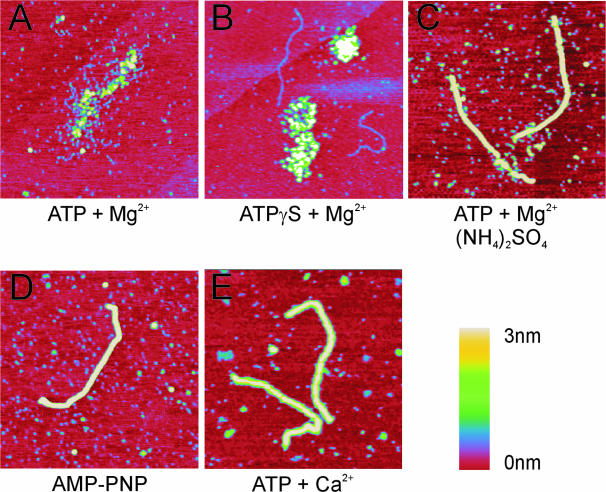
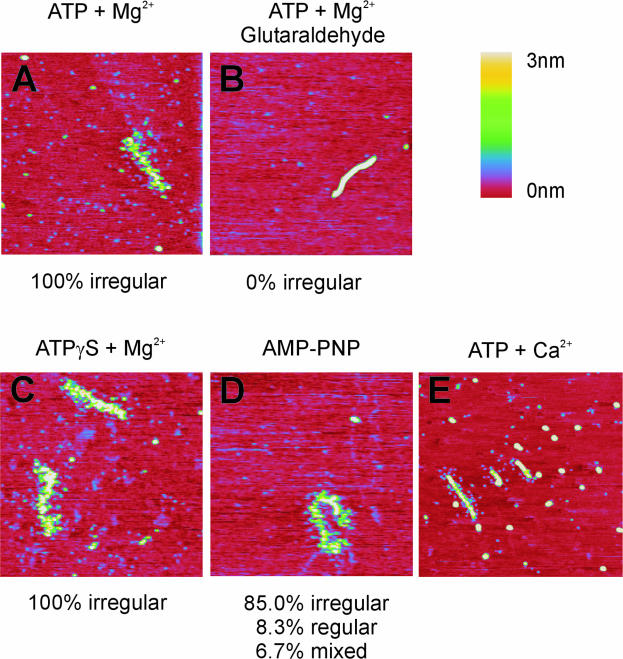
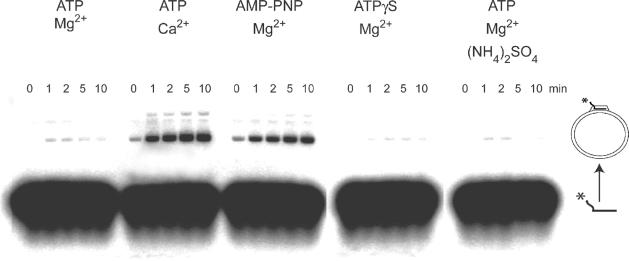
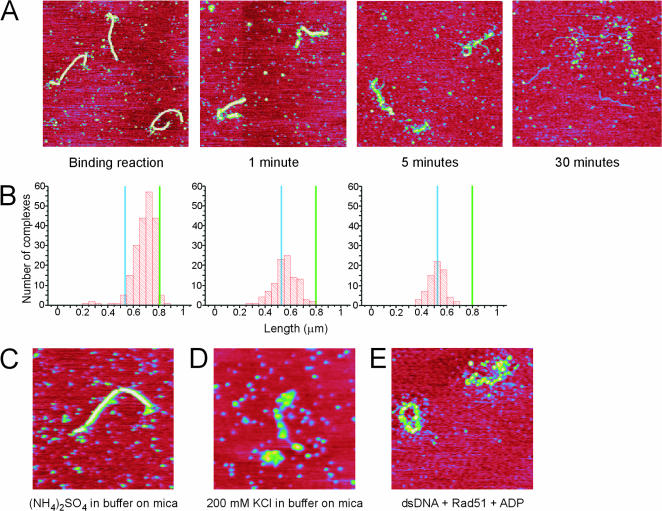
Recombinase proteins assembled into helical filaments on DNA are believed to be the catalytic core of homologous recombination. The assembly, disassembly and dynamic rearrangements of this structure must drive the DNA strand exchange reactions of homologous recombination. The sensitivity of eukaryotic recombinase activity to reaction conditions in vitro suggests that the status of bound nucleotide cofactors is important for function and possibly for filament structure. We analyzed nucleoprotein filaments formed by the human recombinase Rad51 in a variety of conditions on double-stranded and single-stranded DNA by scanning force microscopy. Regular filaments with extended double-stranded DNA correlated with active in vitro recombination, possibly due to stabilizing the DNA products of these assays. Though filaments formed readily on single-stranded DNA, they were very rarely regular structures. The irregular structure of filaments on single-stranded DNA suggests that Rad51 monomers are dynamic in filaments and that regular filaments are transient. Indeed, single molecule force spectroscopy of Rad51 filament assembly and disassembly in magnetic tweezers revealed protein association and disassociation from many points along the DNA, with kinetics different from those of RecA. The dynamic rearrangements of proteins and DNA within Rad51 nucleoprotein filaments could be key events driving strand exchange in homologous recombination.
Figures


References
-
- DiCapua E., Engel A., Stasiak A., Kooller T. Characterization of complexes between recA protein and duplex DNA by electron microscopy. J. Mol. Biol. 1982;157:87–103. - PubMed
-
- Dunn K., Chrysogelos S., Griffith J. Electron microscopic visualization of RecA-DNA filaments: evidence for cyclic extension of duplex DNA. Cell. 1982;28:757–765. - PubMed
-
- Flory J., Radding C.M. Visualization of RecA protein and its association with DNA: a priming effect of single-strand binding protein. Cell. 1982;28:747–756. - PubMed
-
- Yu X., VanLoock M.S., Yang S., Reese J.T., Egelman E.H. What is the structure of the RecA-DNA filament? Curr. Protein Pept. Sci. 2004;5:73–79. - PubMed
-
- Liu Y., Stasiak A.Z., Masson J.Y., McIlwraith M.J., Stasiak A., West S.C. Conformational changes modulate the activity of human RAD51 protein. J. Mol. Biol. 2004;337:817–827. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
