

Correlation of structural elements and infectivity of the HET-s prion
- PMID: 15944710
- PMCID: PMC1567094
- DOI: 10.1038/nature03793
Correlation of structural elements and infectivity of the HET-s prion
Abstract
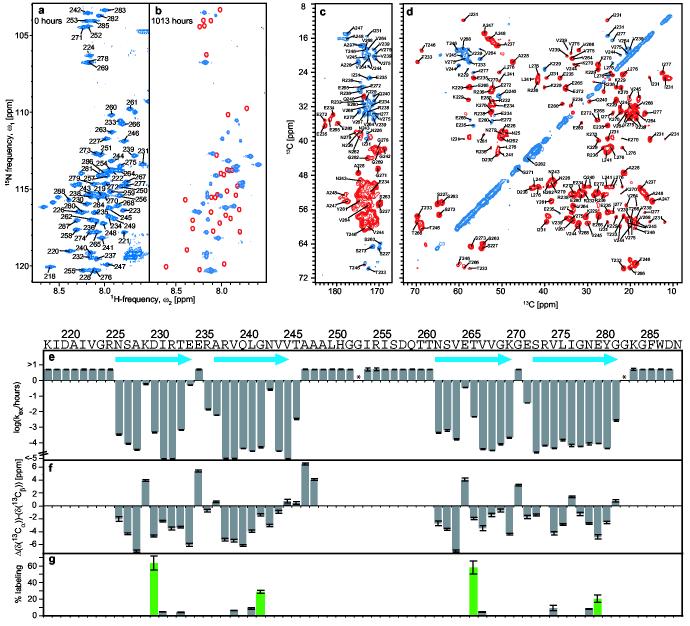
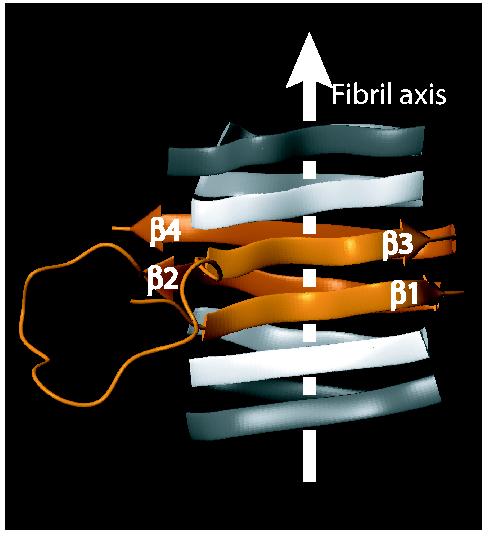
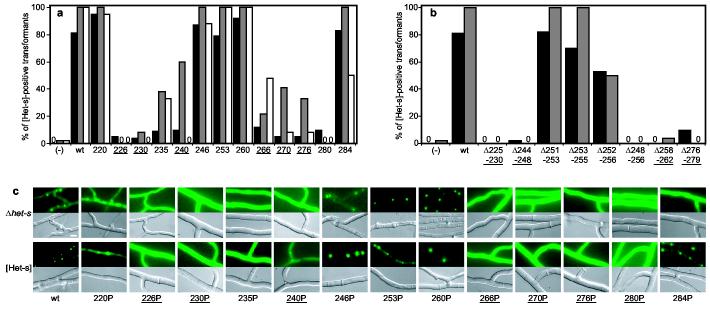
Prions are believed to be infectious, self-propagating polymers of otherwise soluble, host-encoded proteins. This concept is now strongly supported by the recent findings that amyloid fibrils of recombinant prion proteins from yeast, Podospora anserina and mammals can induce prion phenotypes in the corresponding hosts. However, the structural basis of prion infectivity remains largely elusive because acquisition of atomic resolution structural properties of amyloid fibrils represents a largely unsolved technical challenge. HET-s, the prion protein of P. anserina, contains a carboxy-terminal prion domain comprising residues 218-289. Amyloid fibrils of HET-s(218-289) are necessary and sufficient for the induction and propagation of prion infectivity. Here, we have used fluorescence studies, quenched hydrogen exchange NMR and solid-state NMR to determine the sequence-specific positions of amyloid fibril secondary structure elements of HET-s(218-289). This approach revealed four beta-strands constituted by two pseudo-repeat sequences, each forming a beta-strand-turn-beta-strand motif. By using a structure-based mutagenesis approach, we show that this conformation is the functional and infectious entity of the HET-s prion. These results correlate distinct structural elements with prion infectivity.
Figures
Comment in
-
Structural biology: prying into prions.Nature. 2005 Jun 9;435(7043):747-9. doi: 10.1038/435747a. Nature. 2005. PMID: 15944684 No abstract available.
References
-
- Alper T, Cramp WA, Haig DA, Clarke MC. Does the agent of scrapie replicate without nucleic acid? Nature. 1967;214:764–6. - PubMed
-
- Prusiner SB. Novel proteinaceous infectious particles cause scrapie. Science. 1982;216:136–44. - PubMed
-
- Sparrer HE, Santoso A, Szoka FC, Jr., Weissman JS. Evidence for the prion hypothesis: induction of the yeast [PSI+] factor by in vitro- converted Sup35 protein. Science. 2000;289:595–9. - PubMed
-
- King CY, Diaz-Avalos R. Protein-only transmission of three yeast prion strains. Nature. 2004;428:319–23. - PubMed
-
- Tanaka M, Chien P, Naber N, Cooke R, Weissman JS. Conformational variations in an infectious protein determine prion strain differences. Nature. 2004;428:323–8. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
