

Migration of human melanoma cells depends on extracellular pH and Na+/H+ exchange
- PMID: 15946960
- PMCID: PMC1474168
- DOI: 10.1113/jphysiol.2005.088344
Migration of human melanoma cells depends on extracellular pH and Na+/H+ exchange
Abstract
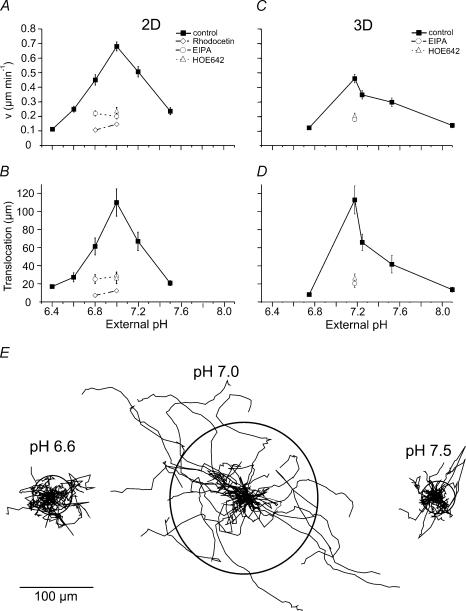
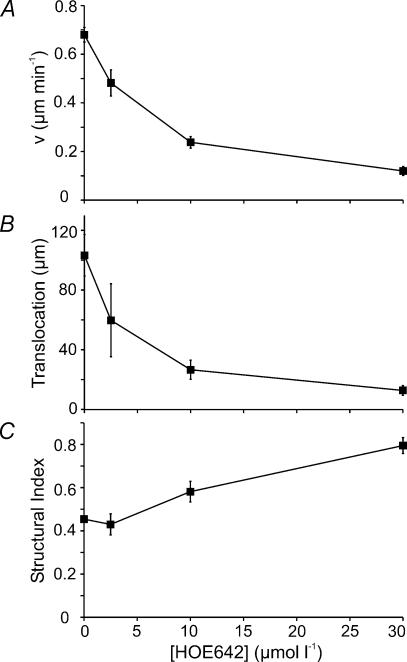
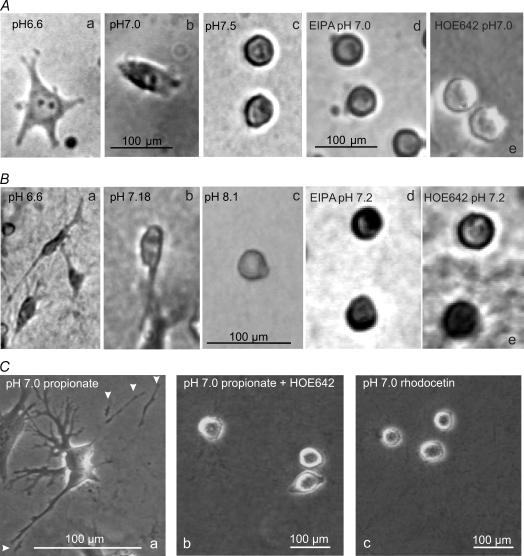
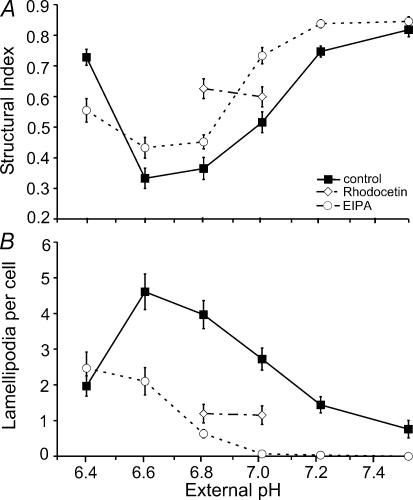
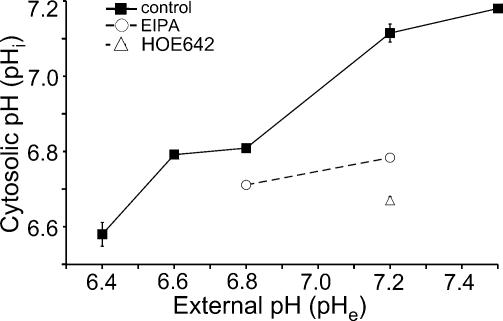
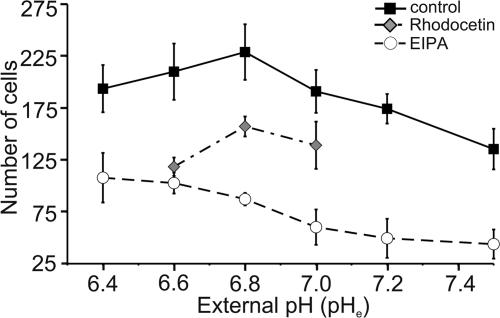
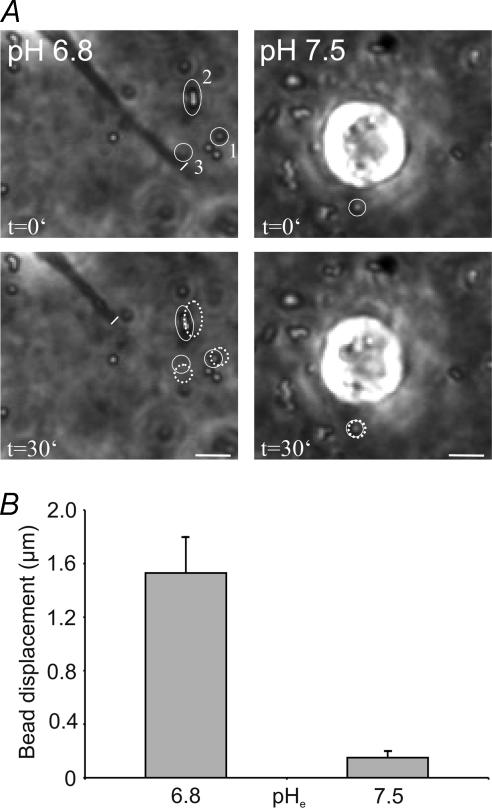
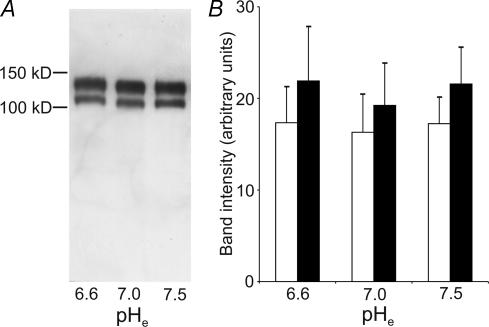
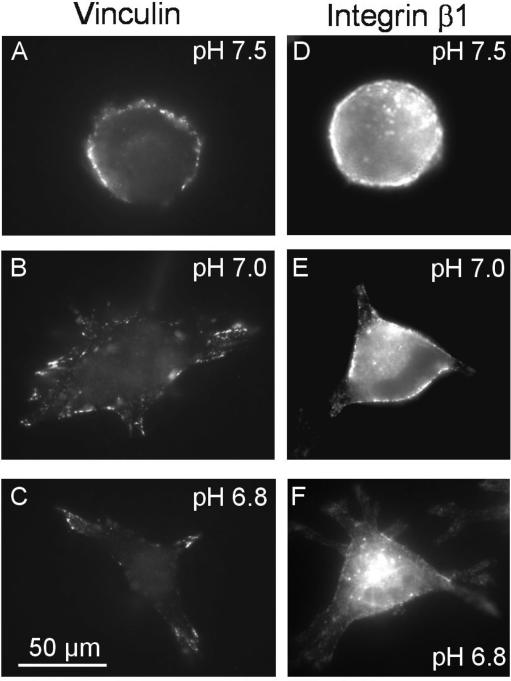
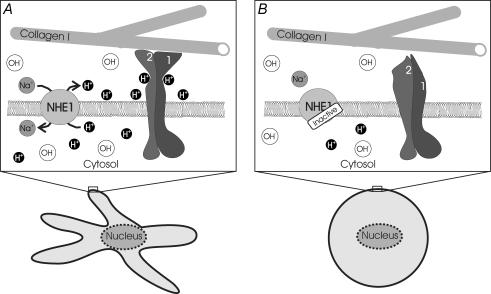
Their glycolytic metabolism imposes an increased acid load upon tumour cells. The surplus protons are extruded by the Na+/H+ exchanger (NHE) which causes an extracellular acidification. It is not yet known by what mechanism extracellular pH (pHe) and NHE activity affect tumour cell migration and thus metastasis. We studied the impact of pHe and NHE activity on the motility of human melanoma (MV3) cells. Cells were seeded on/in collagen I matrices. Migration was monitored employing time lapse video microscopy and then quantified as the movement of the cell centre. Intracellular pH (pHi) was measured fluorometrically. Cell-matrix interactions were tested in cell adhesion assays and by the displacement of microbeads inside a collagen matrix. Migration depended on the integrin alpha2beta1. Cells reached their maximum motility at pHe approximately 7.0. They hardly migrated at pHe 6.6 or 7.5, when NHE was inhibited, or when NHE activity was stimulated by loading cells with propionic acid. These procedures also caused characteristic changes in cell morphology and pHi. The changes in pHi, however, did not account for the changes in morphology and migratory behaviour. Migration and morphology more likely correlate with the strength of cell-matrix interactions. Adhesion was the strongest at pHe 6.6. It weakened at basic pHe, upon NHE inhibition, or upon blockage of the integrin alpha2beta1. We propose that pHe and NHE activity affect migration of human melanoma cells by modulating cell-matrix interactions. Migration is hindered when the interaction is too strong (acidic pHe) or too weak (alkaline pHe or NHE inhibition).
Figures
Similar articles
-
pH dependence of melanoma cell migration: protons extruded by NHE1 dominate protons of the bulk solution.J Physiol. 2007 Dec 1;585(Pt 2):351-60. doi: 10.1113/jphysiol.2007.145185. Epub 2007 Oct 4. J Physiol. 2007. PMID: 17916606 Free PMC article.
-
Regulation of intracellular pH in human melanoma: potential therapeutic implications.Mol Cancer Ther. 2002 Jun;1(8):617-28. Mol Cancer Ther. 2002. PMID: 12479222
-
Functional hierarchy of simultaneously expressed adhesion receptors: integrin alpha2beta1 but not CD44 mediates MV3 melanoma cell migration and matrix reorganization within three-dimensional hyaluronan-containing collagen matrices.Mol Biol Cell. 1999 Oct;10(10):3067-79. doi: 10.1091/mbc.10.10.3067. Mol Biol Cell. 1999. PMID: 10512851 Free PMC article.
-
NHE-1: a molecular target for signalling and cell matrix interactions.Connect Tissue Res. 2008;49(3):157-61. doi: 10.1080/03008200802151581. Connect Tissue Res. 2008. PMID: 18661333 Review.
-
Protons extruded by NHE1: digestive or glue?Eur J Cell Biol. 2008 Sep;87(8-9):591-9. doi: 10.1016/j.ejcb.2008.01.007. Epub 2008 Mar 6. Eur J Cell Biol. 2008. PMID: 18328592 Review.
Cited by
-
Role of the Hedgehog Pathway and CAXII in Controlling Melanoma Cell Migration and Invasion in Hypoxia.Cancers (Basel). 2022 Sep 29;14(19):4776. doi: 10.3390/cancers14194776. Cancers (Basel). 2022. PMID: 36230699 Free PMC article.
-
Cells move when ions and water flow.Pflugers Arch. 2007 Jan;453(4):421-32. doi: 10.1007/s00424-006-0138-6. Epub 2006 Oct 5. Pflugers Arch. 2007. PMID: 17021798 Review.
-
Synergistic activation of genes promoting invasiveness by dual deprivation in oxygen and nutrients.Int J Exp Pathol. 2023 Apr;104(2):64-75. doi: 10.1111/iep.12464. Epub 2023 Jan 24. Int J Exp Pathol. 2023. PMID: 36694990 Free PMC article.
-
Acid-base homeostasis orchestrated by NHE1 defines the pancreatic stellate cell phenotype in pancreatic cancer.JCI Insight. 2023 Oct 9;8(19):e170928. doi: 10.1172/jci.insight.170928. JCI Insight. 2023. PMID: 37643024 Free PMC article.
-
Na+-H+ exchanger-1 (NHE1) regulation in kidney proximal tubule.Cell Mol Life Sci. 2015 Jun;72(11):2061-74. doi: 10.1007/s00018-015-1848-8. Epub 2015 Feb 14. Cell Mol Life Sci. 2015. PMID: 25680790 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical