

Circadian rhythms from multiple oscillators: lessons from diverse organisms
- PMID: 15951747
- PMCID: PMC2735866
- DOI: 10.1038/nrg1633
Circadian rhythms from multiple oscillators: lessons from diverse organisms
Abstract
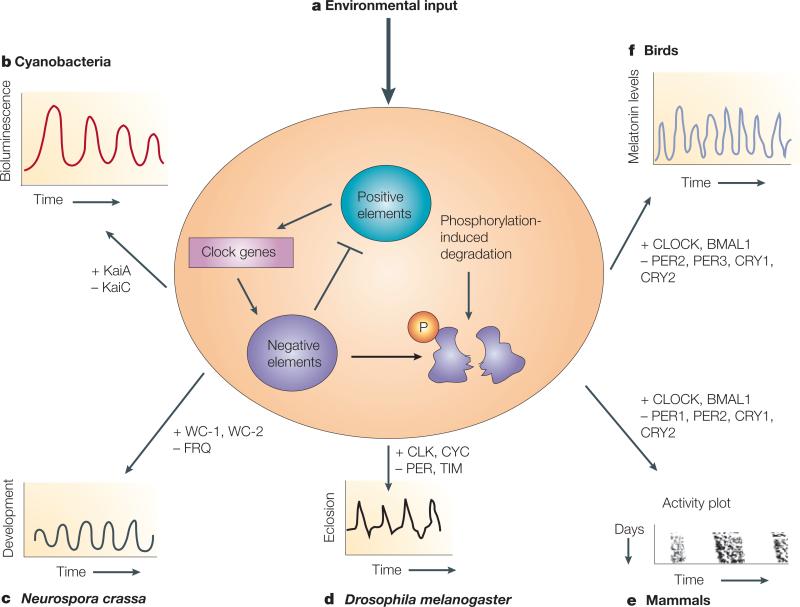
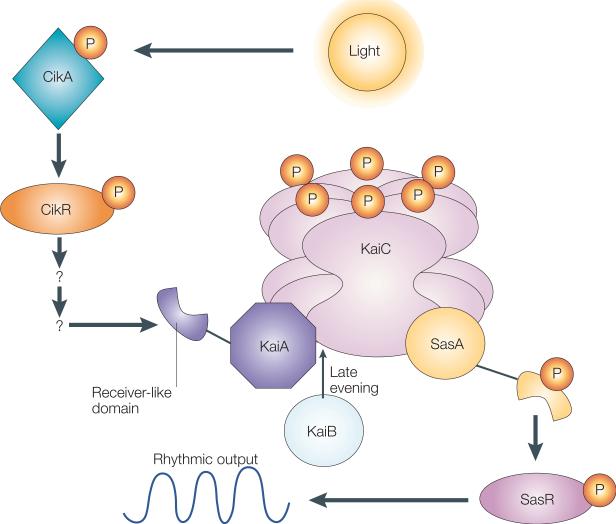
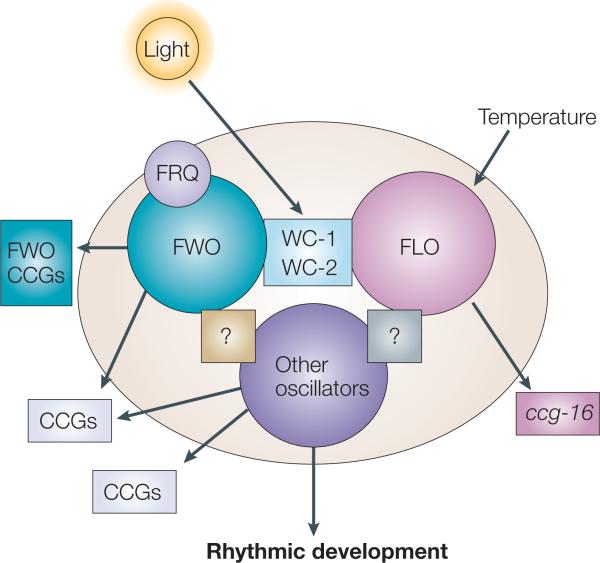
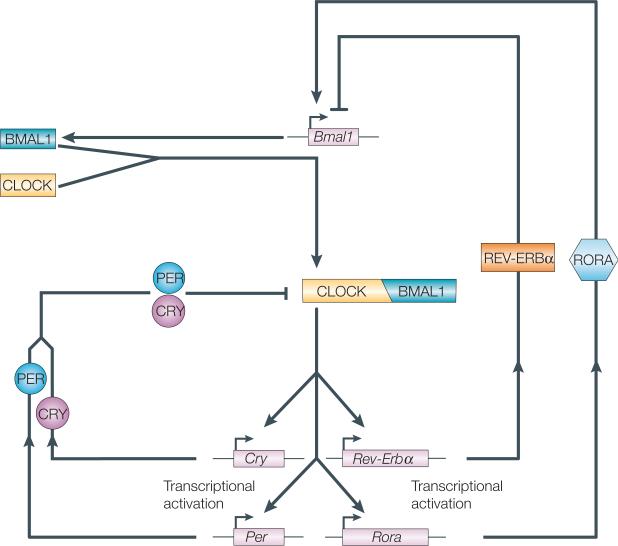
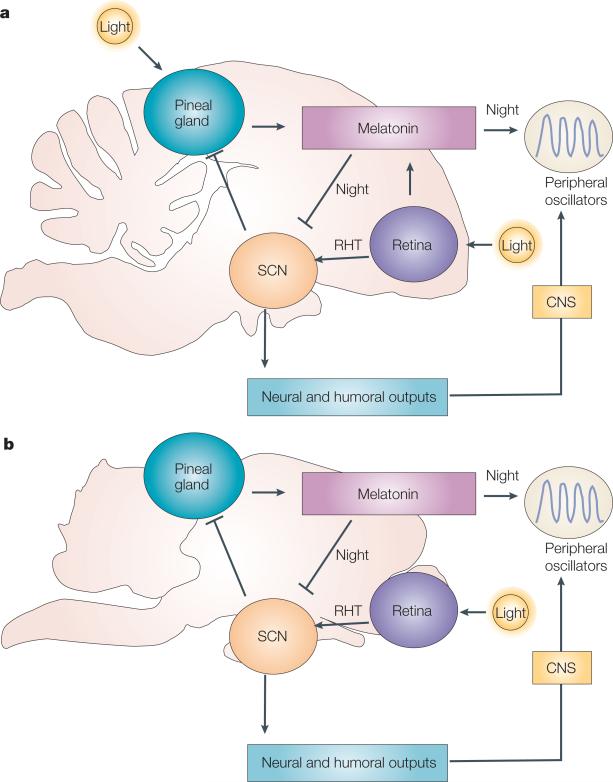
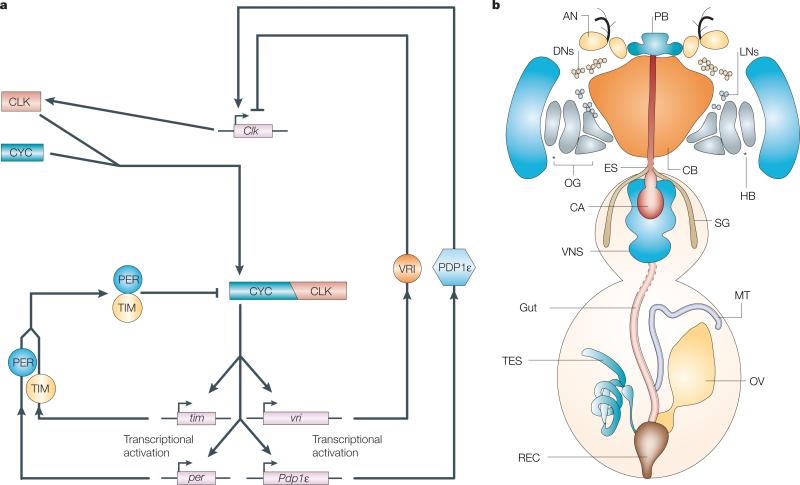
The organization of biological activities into daily cycles is universal in organisms as diverse as cyanobacteria, fungi, algae, plants, flies, birds and man. Comparisons of circadian clocks in unicellular and multicellular organisms using molecular genetics and genomics have provided new insights into the mechanisms and complexity of clock systems. Whereas unicellular organisms require stand-alone clocks that can generate 24-hour rhythms for diverse processes, organisms with differentiated tissues can partition clock function to generate and coordinate different rhythms. In both cases, the temporal coordination of a multi-oscillator system is essential for producing robust circadian rhythms of gene expression and biological activity.
Figures
References
-
- Reppert SM, Weaver DR. Coordination of circadian timing in mammals. Nature. 2002;418:935–941. - PubMed
-
- Young MW, Kay SA. Time zones: a comparative genetics of circadian clocks. Nature Rev. Genet. 2001;2:702–715. - PubMed
-
- Hardin PE. Transcription regulation within the circadian clock: the E-box and beyond. J. Biol. Rhythms. 2004;19:348–360. - PubMed
-
- Hastings MH, Herzog ED. Clock genes, oscillators, and cellular networks in the suprachiasmatic nuclei. J. Biol. Rhythms. 2004;19:400–413. - PubMed
-
- Dunlap JC, Loros JJ. The Neurospora circadian system. J. Biol. Rhythms. 2004;19:414–424. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
