

doi: 10.1104/pp.104.058362.
Maturation of the ground tissue of the root is regulated by gibberellin and SCARECROW and requires SHORT-ROOT
Affiliations
- PMID: 15955927
- PMCID: PMC1150384
- DOI: 10.1104/pp.104.058362
Item in Clipboard
Maturation of the ground tissue of the root is regulated by gibberellin and SCARECROW and requires SHORT-ROOT
Plant Physiol.
2005 Jun.
No abstract available
Figures
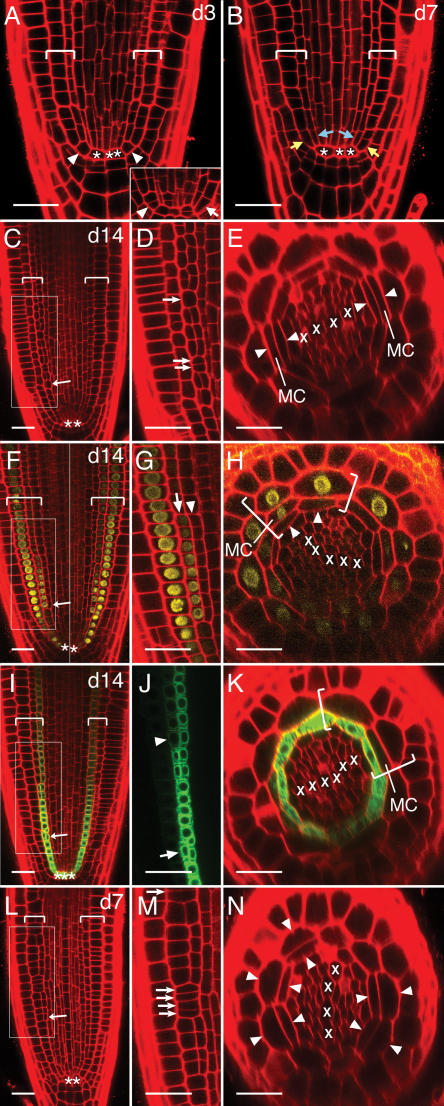
Maturation of the wild-type Arabidopsis primary root and regulation of the process by SCR. A and B, Over time, separate stem cells form for each of the two layers of ground tissue by periclinal division of the C/E stem cell. A, At d3 many C/E stem cells are not divided (arrowheads), but the process is beginning (inset, arrow). B, By d7 almost all roots show separate cortex stem cells (yellow arrows) and endodermis stem cells (blue arrows). C to K, By d14 middle cortex is beginning to form. Areas boxed in C, F, and I are shown magnified in D, G, and J, respectively. C to E, The first cells of the middle cortex arise from periclinal division of endodermal-layer cells that are not near the QC (C, arrow) and that are adjacent to the xylem poles (E, arrowheads). At this stage, within a cell file, divided cells are frequently interspersed with undivided cells (D, arrows). F to H, Cells of the middle cortex rapidly take on cortical identity. F, Composite image of two focal planes of the same root. The cortex-specific marker pCo2∷YFPH2B is expressed de novo in all outer derivatives of divided endodermal-layer cells (G, arrow). pCo2∷YFPH2B is induced in middle-cortex cells but not in adjacent endodermal cells (G and H, arrowheads). I to K, Cells of the middle cortex lose endodermal identity. The endodermis-specific marker SCR∷green fluorescent protein is eventually downregulated in middle-cortex cells that are distant from the QC (J, compare arrowhead to arrow; K, MC). K, Shows a different root than in I and J. L–N, SCR is not required for the divisions that normally form middle cortex. SCR may, in fact, normally act to repress these divisions in both time and space. At d7 divisions of ground tissue cells in the scr-5 null mutant resemble middle-cortex-forming divisions in wild type with respect to their distance from the QC (compare L to C, arrows) and their discontinuous pattern within a cell file (compare M to D, arrows). In a scr null mutant the first divisions occur at an earlier time point and in a position different from that in wild type (compare N to E, arrowheads relative to Xs). Longitudinal (A–D, F, G, I, J, L, M) and transverse (E, H, K, N) optical sections of the apical meristematic zone. Brackets, Ground tissue; asterisk, QC cell; X, xylem progenitor. Scale bars, 25 microns.

Effect of inhibition of GA biosynthesis on timing and extent of middle-cortex formation in wild type and its stronger effect in scr. Longitudinal optical sections of the primary root apical meristem. A to C, Wild type at d7. D to F, scr-5 at d3. A, Mock-treated wild-type seedlings show no formation of middle cortex at d7. Inhibition of GA biosynthesis by growth on 2 μm pac (B) and by removal of copalyl diphosphate synthase function (i.e. introduction of the ga1-3 mutation into Col) (C) each lead to premature initiation of middle-cortex formation (arrowheads and joined arrows). D, Middle-cortex-forming divisions are just beginning at d3 in mock-treated scr-5 seedlings (arrowhead). Growth on 0.5 μm pac (E) and addition of the ga1-3 mutation (F) each lead to more extensive middle-cortex formation in a greater proportion of roots (arrowhead and joined arrows). Brackets, Ground tissue. Scale bars, 25 microns.
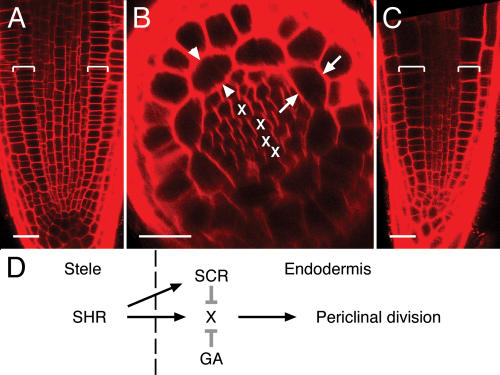
Lack of effect of inhibition of GA biosynthesis on the shr mutant and a model. Longitudinal (A and C) and transverse (B) optical sections of the primary root apical meristem of shr-2 at d7. shr-2 shows no middle-cortex formation, even when grown on 2 μm pac (A and B) or when combined with the ga1-3 mutation (C). Divisions occur neither adjacent to the phloem poles (B, arrows) nor at the ends of the xylem axis (B, arrowheads). D, One possible model for the role of SHR, SCR, and GA in the developmental transition of initiation of middle-cortex formation by periclinal divisions of endodermal cells. The divisions require X function in the endodermis, which, in turn, requires SHR (which originates in the innermost tissue of the root, the stele) but does not require SCR. SCR and GA both independently inhibit X function (grey T lines) to varying degrees relative to each other at different stages of root maturation. Brackets, Ground tissue. X, Xylem progenitor. Scale bars, 25 microns.
References
-
- Barley R, Waites R (2002) Plant meristems: the interplay of KNOX and gibberellins. Curr Biol 12: R696–R698 - PubMed
-
- Baum SF, Dubrovsky JG, Rost TL (2002) Apical organization and maturation of the cortex and vascular cylinder in Arabidopsis thaliana (Brassicaceae) roots. Am J Bot 89: 908–920 - PubMed
-
- Benfey PN, Linstead PJ, Roberts K, Schiefelbein JW, Hauser M-T, Aeschbacher RA (1993) Root development in Arabidopsis: four mutants with dramatically altered root morphogenesis. Development 119: 57–70 - PubMed
-
- Benfey PN, Scheres B (2000) Root development. Curr Biol 10: R813–R815 - PubMed
-
- Doerner P (2003) Plant meristems: a merry-go-round of signals. Curr Biol 13: R368–R374 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
