

Adult myogenesis in Drosophila melanogaster can proceed independently of myocyte enhancer factor-2
- PMID: 15956678
- PMCID: PMC1449755
- DOI: 10.1534/genetics.105.041749
Adult myogenesis in Drosophila melanogaster can proceed independently of myocyte enhancer factor-2
Abstract
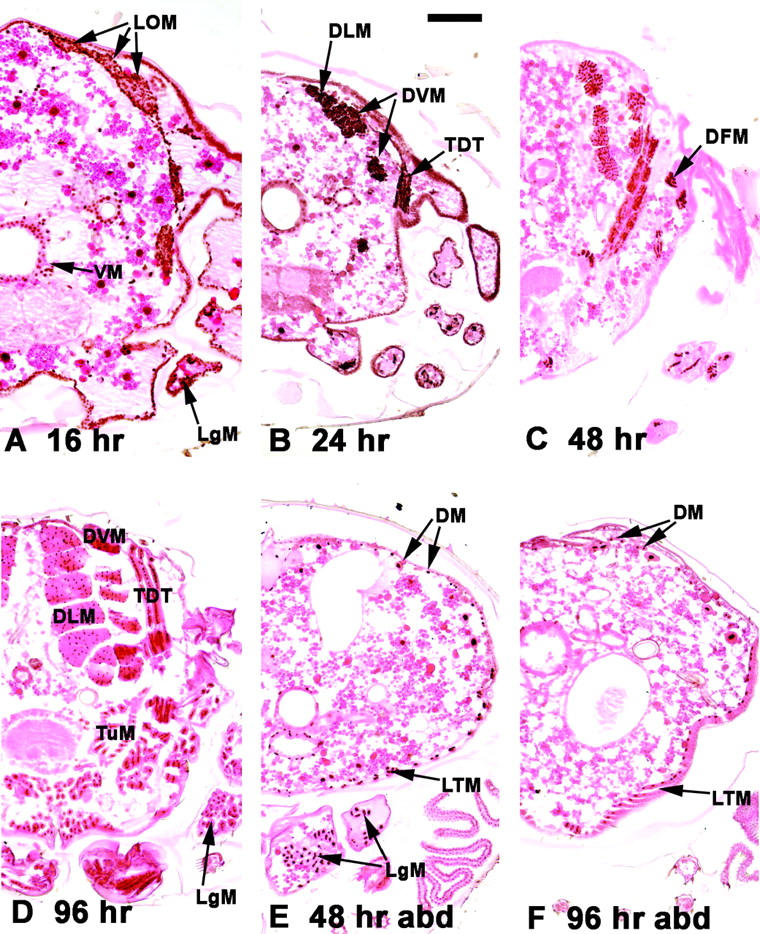
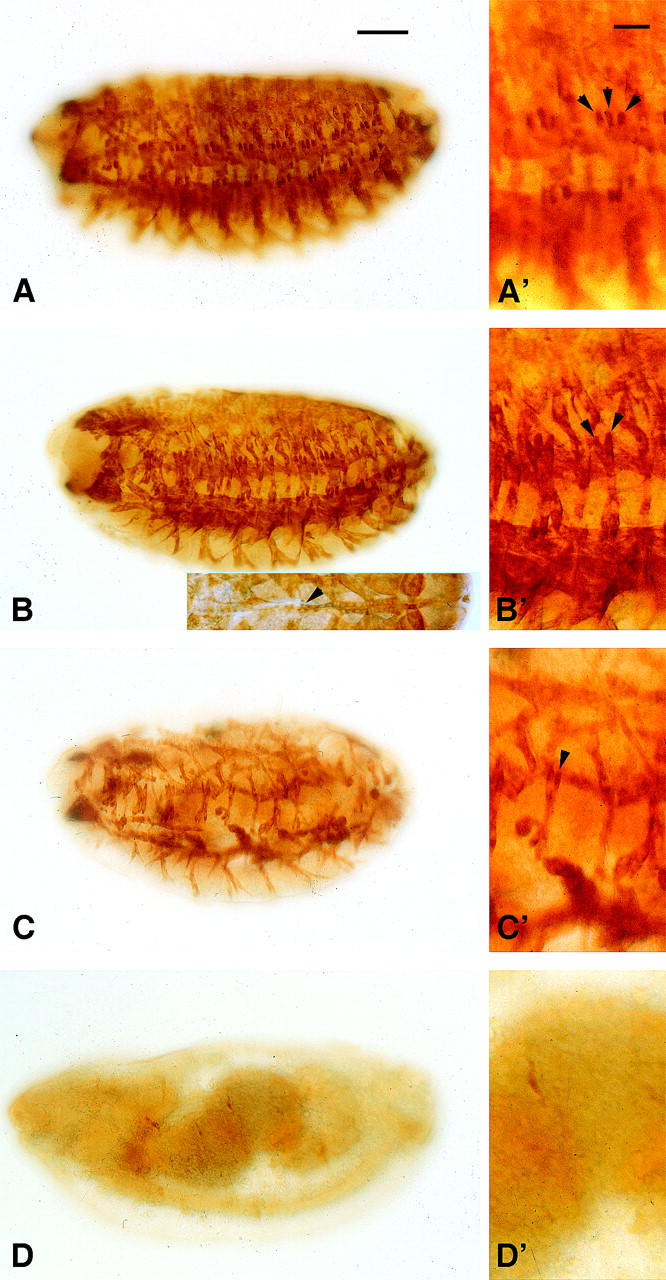
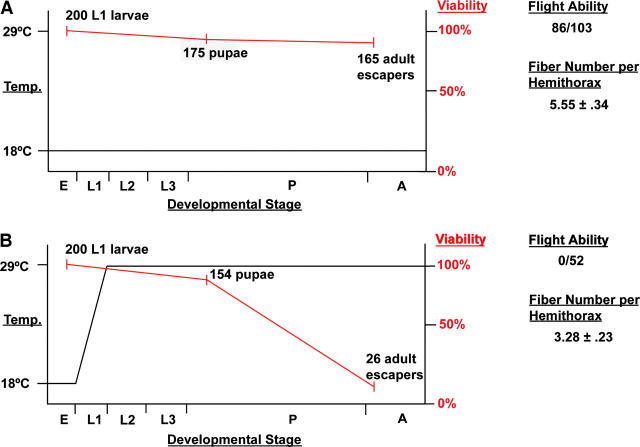
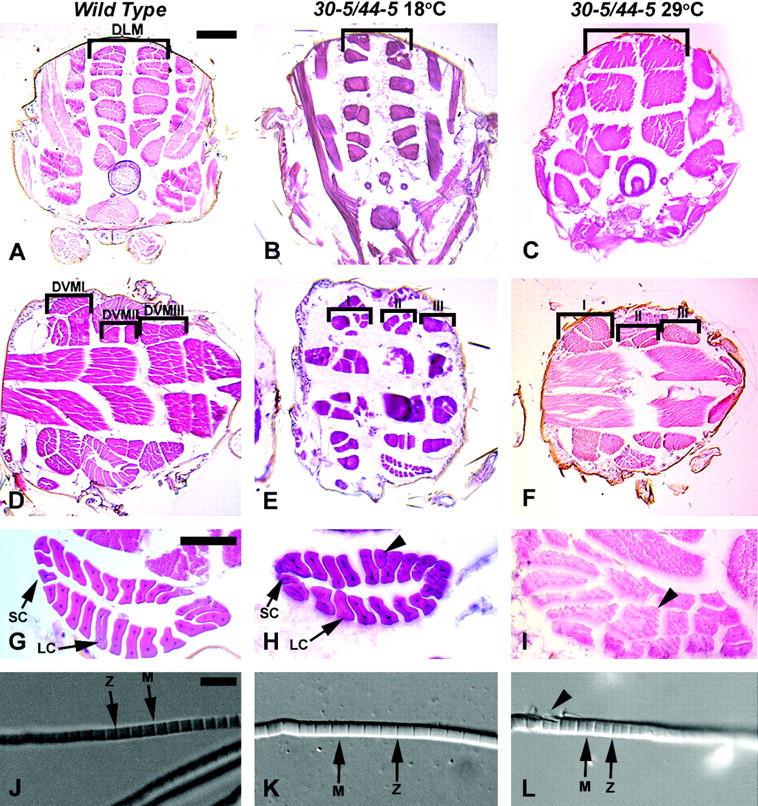
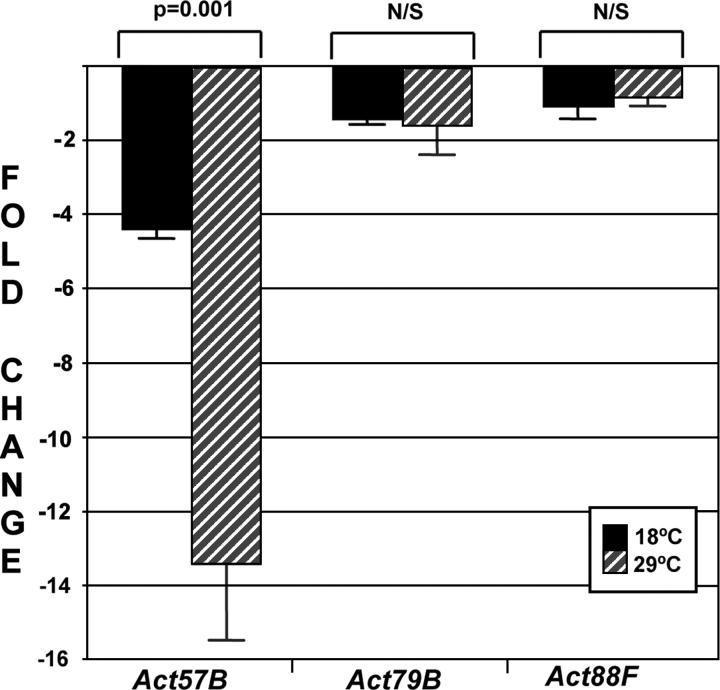
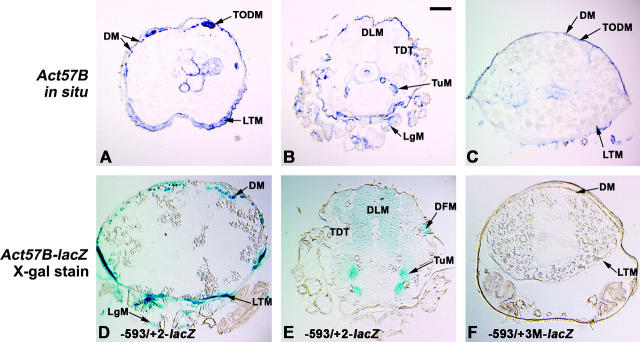
Myocyte enhancer factor-2 (MEF2) is a transcription factor that is necessary for embryonic muscle development in Drosophila and vertebrates; however, whether this factor is required during later muscle development remains largely unknown. Using heteroallelic combinations of different Mef2 mutant alleles, we isolated and characterized a temperature-sensitive combination. Through temperature-shift experiments, we obtained adult animals that were lacking proper MEF2 function. Many of these individuals died as mature pupae, and those that eclosed showed poor locomotion and an inability to fly. Histological analysis of these animals revealed a requirement for MEF2 in skeletal muscle patterning, although these animals had strikingly normal amounts of muscle tissue. Using quantitative polymerase chain reaction, we determined that expression of the MEF2-regulated actin gene Act57B was severely reduced in these animals. By contrast myofibrillar actin genes unique to the adult stage were only mildly affected. Since MEF2 mutant adults were still capable of forming muscle tissue, we conclude that MEF2 is required for the expression of only a subset of muscle structural genes in the adult. These results indicate that additional muscle-specific factors function to control the myogenesis of complex and diverse muscle in the adult.
Figures
References
-
- Anant, S., S. Roy and K. Vijayraghavan, 1998. Twist and Notch negatively regulate adult muscle differentiation in Drosophila. Development 125: 1361–1369. - PubMed
-
- Andres, V., M. Cervera and V. Mahdavi, 1995. Determination of the consensus binding site for MEF2 expressed in muscle and brain reveals tissue-specific sequence constraints. J. Biol. Chem. 270: 23246–23249. - PubMed
-
- Arredondo, J. J., R. M. Ferreres, M. Maroto, R. M. Cripps, R. Marco et al., 2001. Control of Drosophila Paramyosin/Miniparamyosin gene expression: differential regulatory mechanisms for muscle-specific transcription. J. Biol. Chem. 276: 8278–8287. - PubMed
-
- Ashburner, M., 1989 Drosophila: A Laboratory Handbook. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY.
-
- Bagni, C., S. Bray, J. A. Gogos, F. C. Kafatos and T. Hsu, 2002. The Drosophila zinc finger transcription factor CF2 is a myogenic marker downstream of MEF2 during muscle development. Mech. Dev. 117: 265–268. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
