

Cell cycle-dependent dynamics and regulation of mitotic kinesins in Drosophila S2 cells
- PMID: 15958489
- PMCID: PMC1182325
- DOI: 10.1091/mbc.e05-02-0118
Cell cycle-dependent dynamics and regulation of mitotic kinesins in Drosophila S2 cells
Abstract
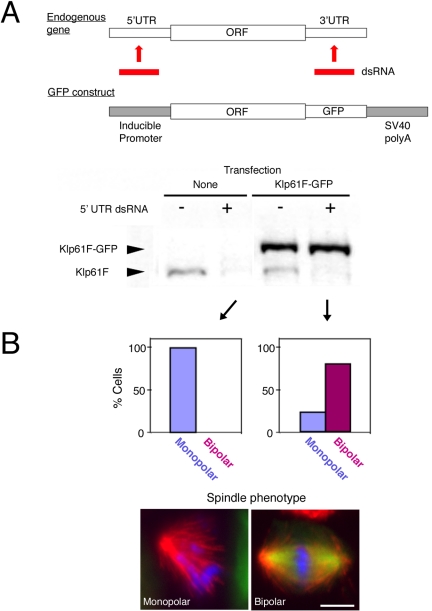
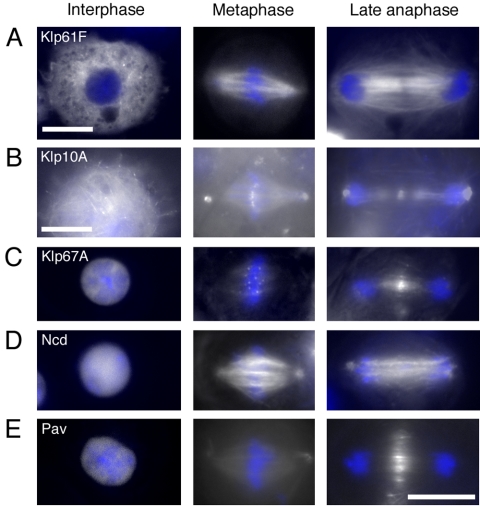
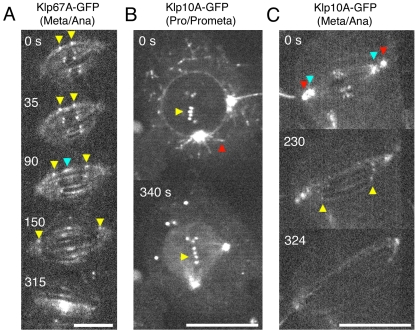
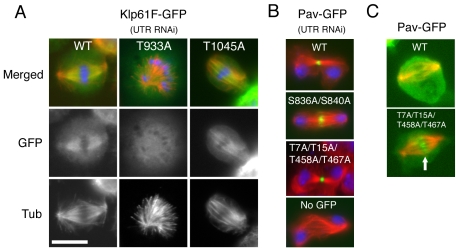
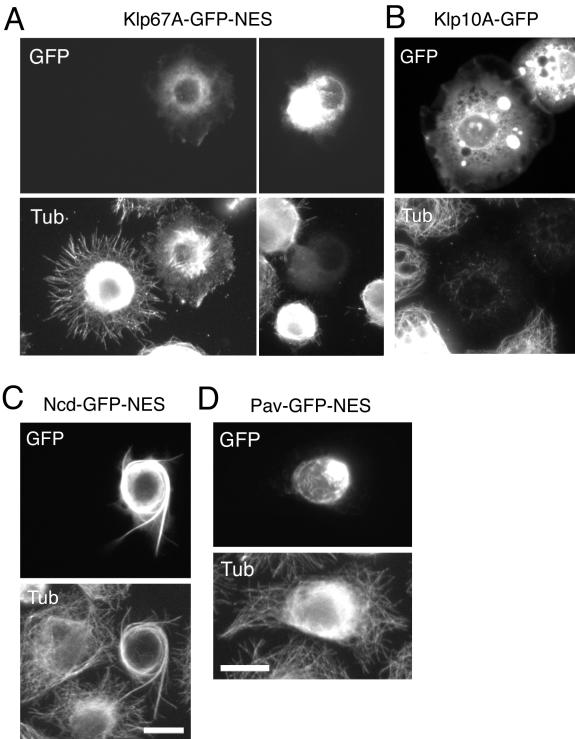
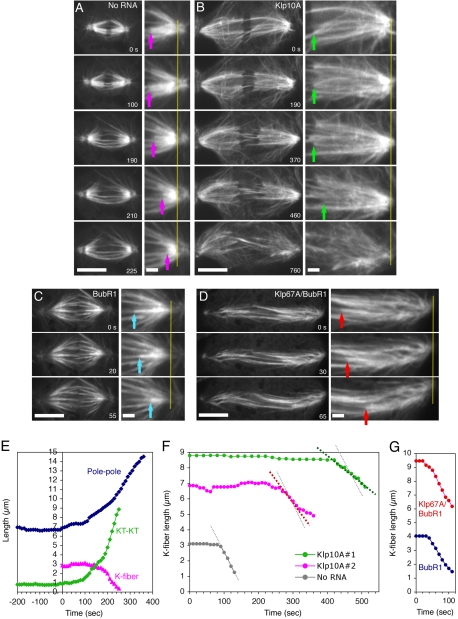
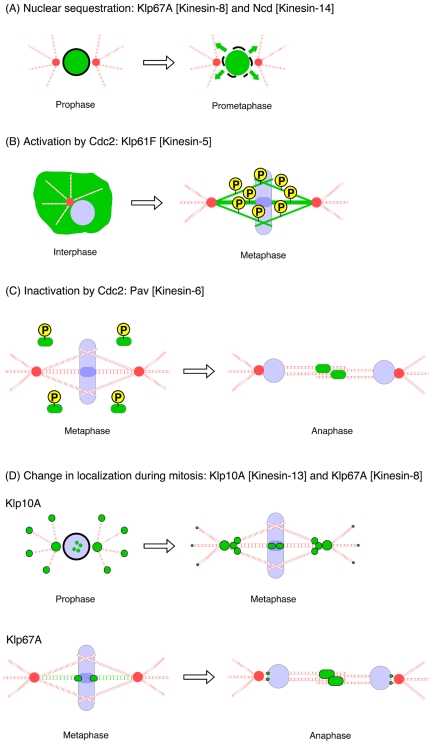
Constructing a mitotic spindle requires the coordinated actions of several kinesin motor proteins. Here, we have visualized the dynamics of five green fluorescent protein (GFP)-tagged mitotic kinesins (class 5, 6, 8, 13, and 14) in live Drosophila Schneider cell line (S2), after first demonstrating that the GFP-tag does not interfere with the mitotic functions of these kinesins using an RNA interference (RNAi)-based rescue strategy. Class 8 (Klp67A) and class 14 (Ncd) kinesin are sequestered in an active form in the nucleus during interphase and engage their microtubule targets upon nuclear envelope breakdown (NEB). Relocalization of Klp67A to the cytoplasm using a nuclear export signal resulted in the disassembly of the interphase microtubule array, providing support for the hypothesis that this kinesin class possesses microtubule-destabilizing activity. The interactions of Kinesin-5 (Klp61F) and -6 (Pavarotti) with microtubules, on the other hand, are activated and inactivated by Cdc2 phosphorylation, respectively, as shown by examining localization after mutating Cdc2 consensus sites. The actions of microtubule-destabilizing kinesins (class 8 and 13 [Klp10A]) seem to be controlled by cell cycle-dependent changes in their localizations. Klp10A, concentrated on microtubule plus ends in interphase and prophase, relocalizes to centromeres and spindle poles upon NEB and remains at these sites throughout anaphase. Consistent with this localization, RNAi analysis showed that this kinesin contributes to chromosome-to-pole movement during anaphase A. Klp67A also becomes kinetochore associated upon NEB, but the majority of the population relocalizes to the central spindle by the timing of anaphase A onset, consistent with our RNAi result showing no effect of depleting this motor on anaphase A. These results reveal a diverse spectrum of regulatory mechanisms for controlling the localization and function of five mitotic kinesins at different stages of the cell cycle.
Figures
References
-
- Blangy, A., Lane, H. A., d'Herin, P., Harper, M., Kress, M., and Nigg, E. A. (1995). Phosphorylation by p34cdc2 regulates spindle association of human Eg5, a kinesin-related motor essential for bipolar spindle formation in vivo. Cell 83, 1159-1169. - PubMed
-
- Compton, D. A. (2000). Spindle assembly in animal cells. Annu. Rev. Biochem. 69, 95-114. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
