

Immunodetection of disease-associated mutant PrP, which accelerates disease in GSS transgenic mice
- PMID: 15962001
- PMCID: PMC1173157
- DOI: 10.1038/sj.emboj.7600717
Immunodetection of disease-associated mutant PrP, which accelerates disease in GSS transgenic mice
Abstract
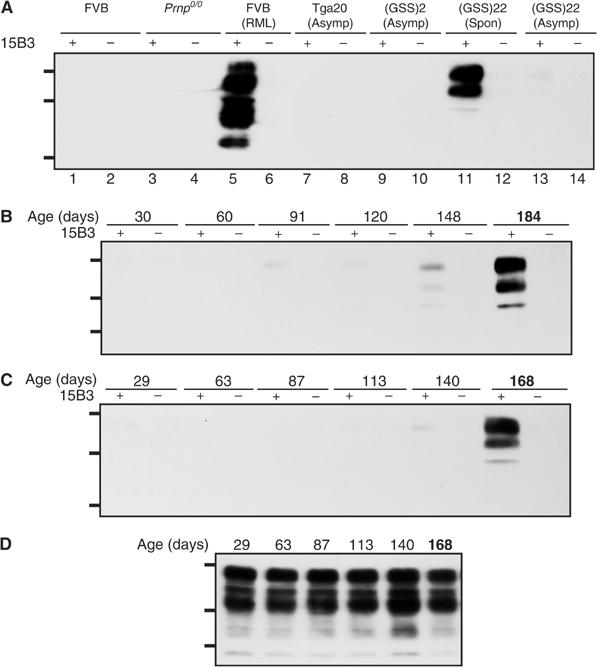
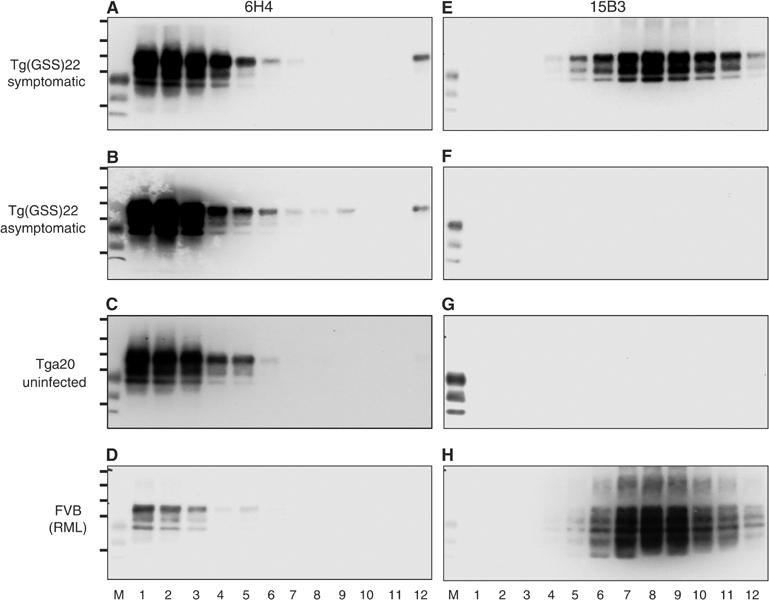
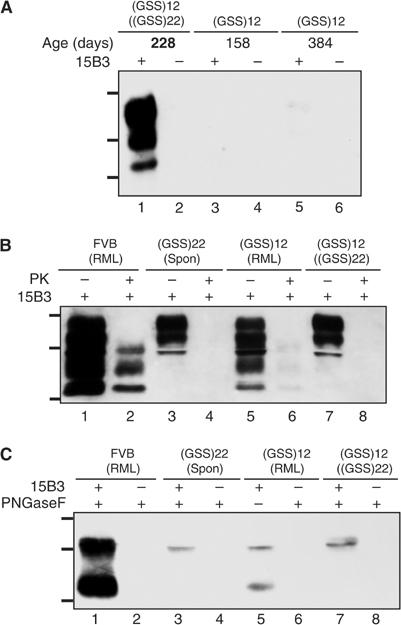
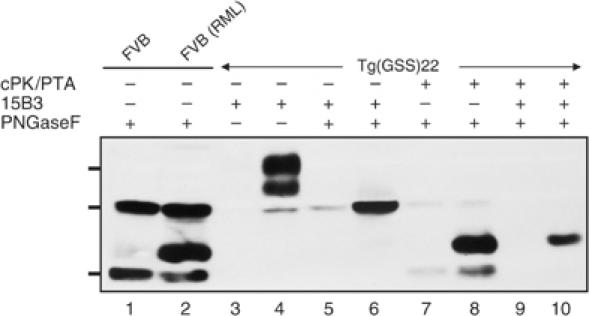
The absence of infectivity-associated, protease-resistant prion protein (PrP(Sc)) in the brains of spontaneously sick transgenic (Tg) mice overexpressing PrP linked to Gerstmann-Sträussler Scheinker syndrome, and the failure of gene-targeted mice expressing such PrP to develop disease spontaneously, challenged the concept that mutant PrP expression led to spontaneous prion production. Here, we demonstrate that disease in overexpressor Tg mice is associated with accumulation of protease-sensitive aggregates of mutant PrP that can be immunoprecipitated by the PrP(Sc)-specific monoclonal antibody designated 15B3. Whereas Tg mice expressing multiple transgenes exhibited accelerated disease when inoculated with disease-associated mutant PrP, Tg mice expressing mutant PrP at low levels failed to develop disease either spontaneously or following inoculation. These studies indicate that inoculated mutant PrP from diseased mice promotes the aggregation and accumulation of pre-existing pathological forms of mutant PrP produced as a result of transgene overexpression. Thus, while pathological mutant PrP possesses a subset of PrP(Sc) characteristics, we now show that the attribute of prion transmission suggested by previous studies is more accurately characterized as disease acceleration.
Figures
References
-
- Brown P, Gibbs CJ Jr, Rodgers-Johnson P, Asher DM, Sulima MP, Bacote A, Goldfarb LG, Gajdusek DC (1994) Human spongiform encephalopathy: the National Institutes of Health series of 300 cases of experimentally transmitted disease. Ann Neurol 35: 513–529 - PubMed
-
- Chandler RL (1961) Encephalopathy in mice produced by inoculation with scrapie brain material. Lancet 1: 1378–1379 - PubMed
-
- Chen SG, Teplow DB, Parchi P, Teller JK, Gambetti P, Autilio-Gambetti L (1995) Truncated forms of the human prion protein in normal brain and in prion diseases. J Biol Chem 270: 19173–19180 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
