

Role of NK-1 neurotransmission in opioid-induced hyperalgesia
- PMID: 15964684
- PMCID: PMC1440305
- DOI: 10.1016/j.pain.2005.04.014
Role of NK-1 neurotransmission in opioid-induced hyperalgesia
Abstract
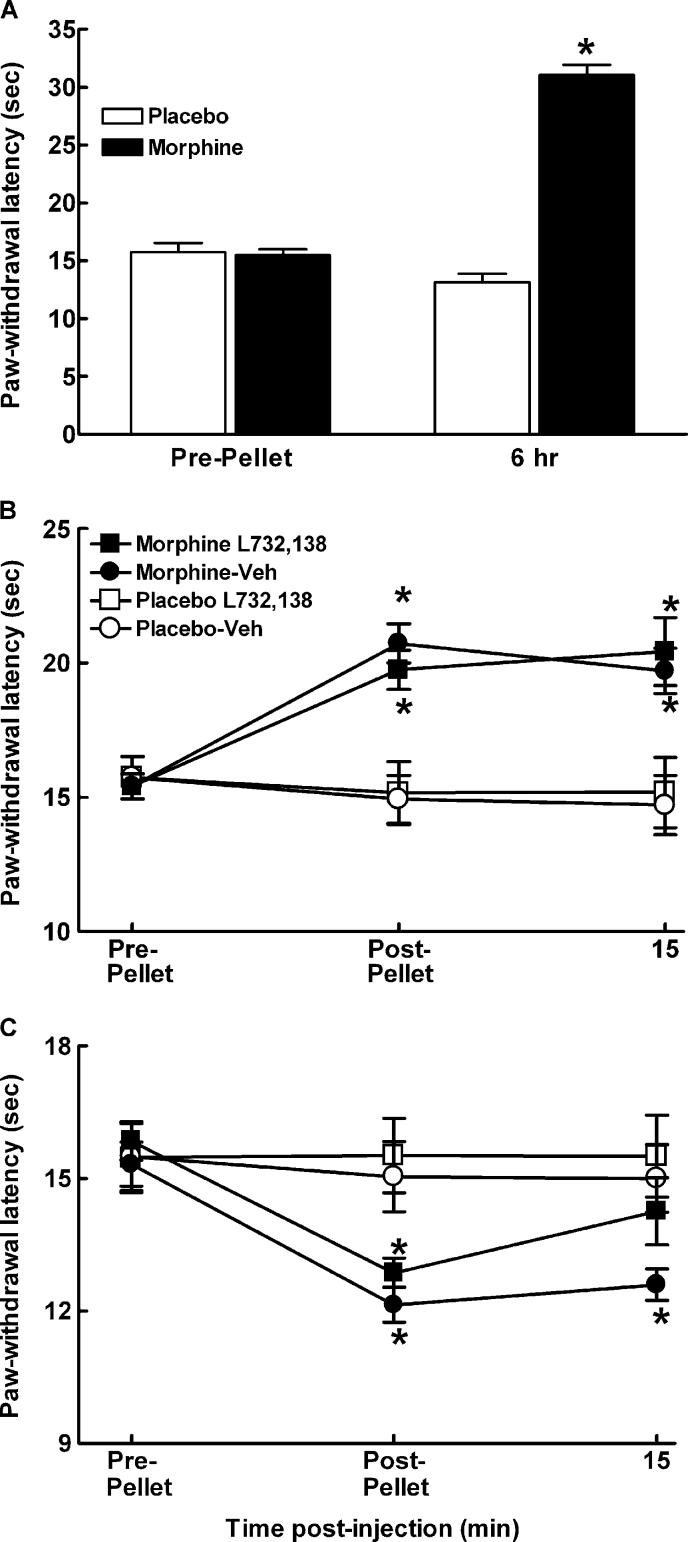
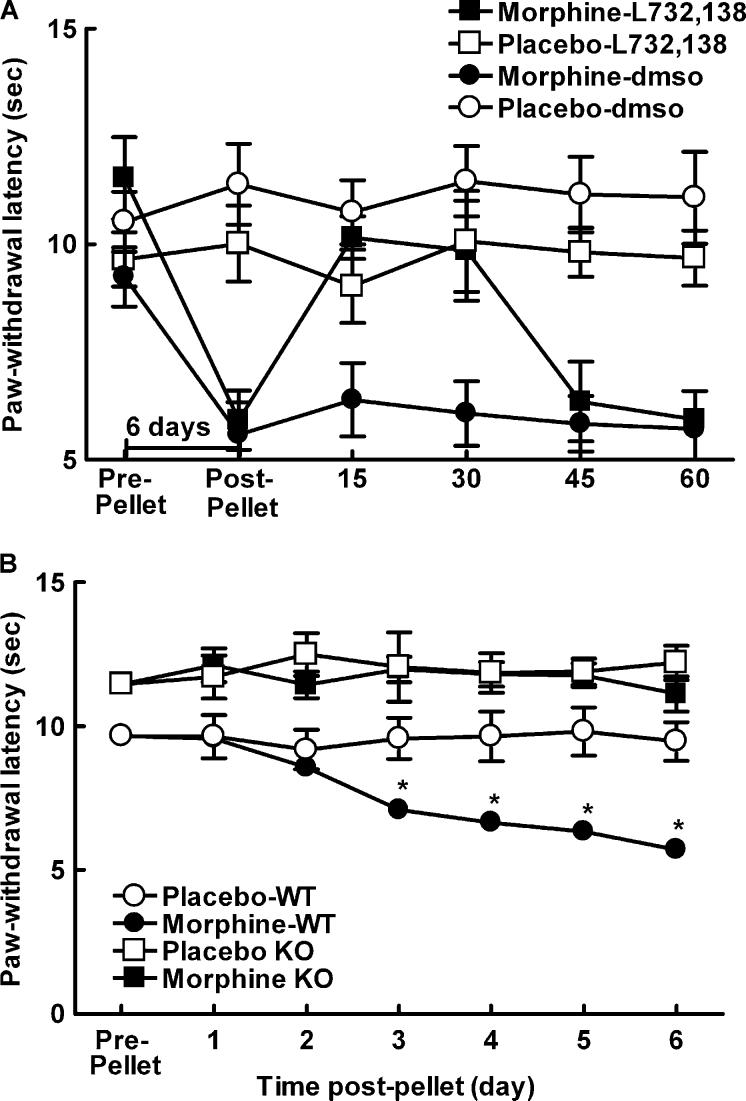
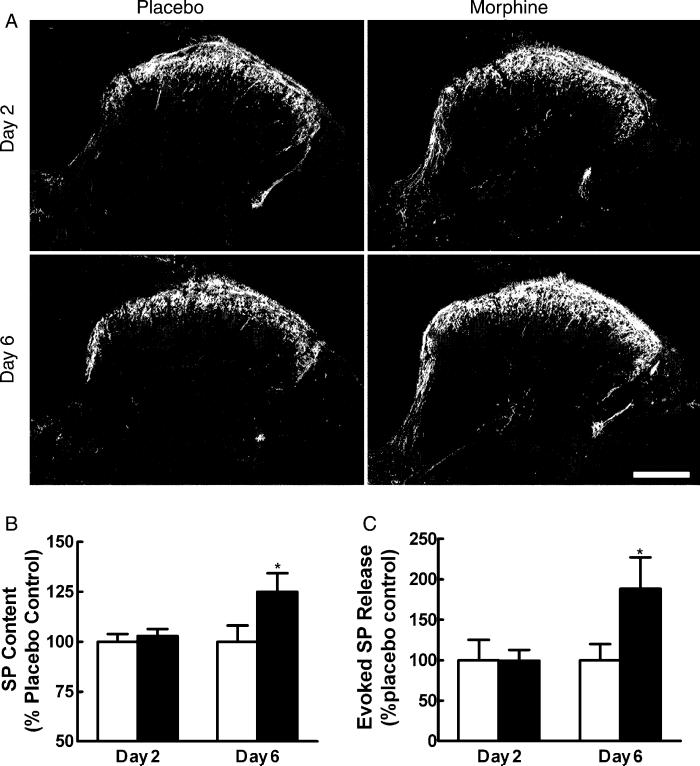
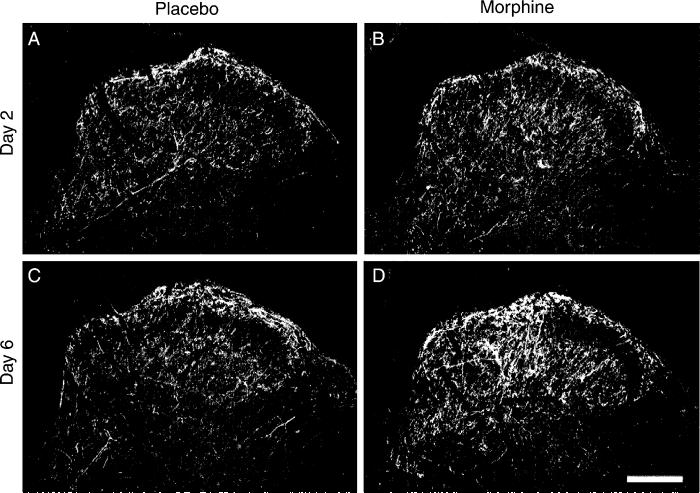
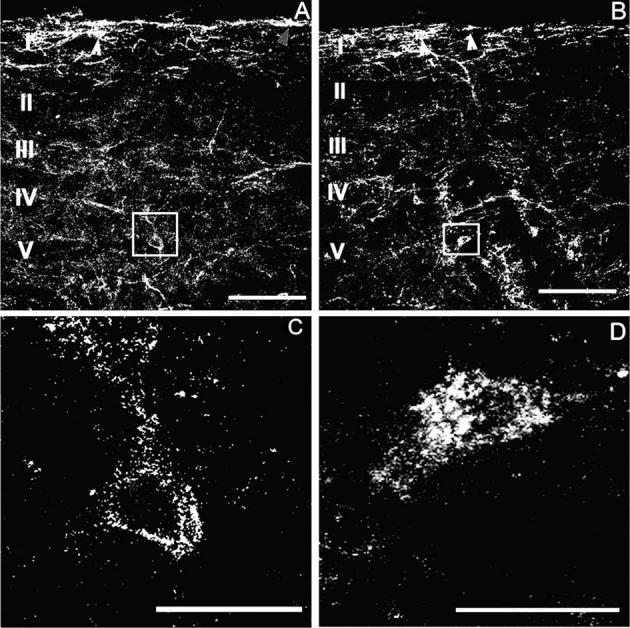
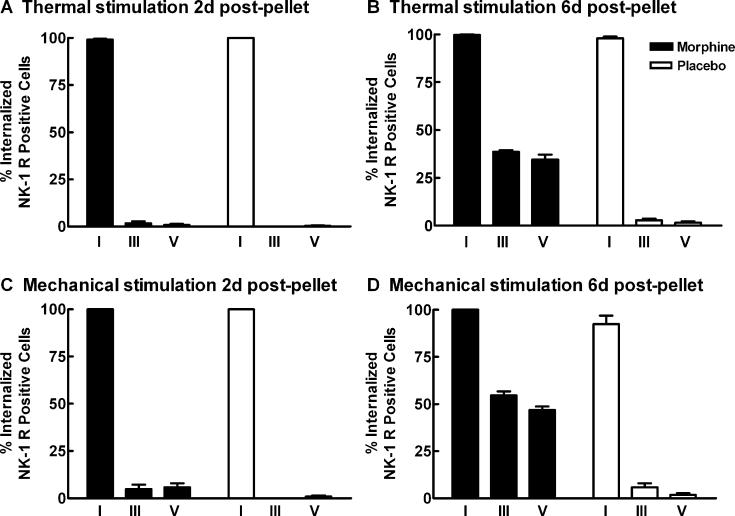
Opiates are among the most important drugs for treatment of moderate to severe pain and prolonged opiate administration is often required to treat chronic pain states. We investigated the neurobiological actions of sustained opiate administration revealing paradoxical pronociceptive adaptations associated with NK-1 receptor function. Sustained morphine delivered over 6 days elicited hyperalgesia in rats and mice during the period of opiate delivery. Sustained morphine administration increased substance P (SP) and NK-1 receptor expression in the spinal dorsal horn. Sustained morphine treatment also enhanced capsaicin-evoked SP release in vitro, and increased internalization of NK-1 receptors in response to noxious stimulation. While NK-1 receptor internalization was observed primarily in the superficial laminae of placebo-treated rats, NK-1 receptor internalization was seen in both superficial and deep lamina of the dorsal horn in morphine-treated animals. Morphine-induced hyperalgesia was reversed by spinal administration of an NK-1 receptor antagonist in rats and mice, and was observed in wildtype (NK-1(+/+)), but not NK-1 receptor knockout (NK-1(-/-)), mice. These data support a critical role for the NK-1 receptor in the expression of sustained morphine-induced hyperalgesia. Additionally, these data indicate that sustained opiate administration induces changes reminiscent of those associated with inflammatory pain. These opiate-induced changes might produce unintended deleterious actions in the course of pain treatment in patients. Understanding of sustained morphine-induced neurochemical changes will help identify approaches that limit the deleterious actions of opiates.
Figures
References
-
- Abbadie C, Brown JL, Mantyh PW, Basbaum AI. Spinal cord substance P receptor immunoreactivity increases in both inflammatory and nerve injury models of persistent pain. Neuroscience. 1996;70:201–9. - PubMed
-
- Cahill CM, Coderre TJ. Attenuation of hyperalgesia in a rat model of neuropathic pain after intrathecal pre- or post-treatment with a neurokinin-1 antagonist. Pain. 2002;95:277–85. - PubMed
-
- Cascieri MA, Macleod AM, Underwood D, Shiao LL, Ber E, Sadowski S, Yu H, Merchant KJ, Swain CJ, Strader CD, Fong TM. Characterization of the interaction of N-acyl-L-tryptophan benzyl ester neurokinin antagonists with the human neurokinin-1 receptor. J Biol Chem. 1994;269:6587–91. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
