

Esf2p, a U3-associated factor required for small-subunit processome assembly and compaction
- PMID: 15964808
- PMCID: PMC1156982
- DOI: 10.1128/MCB.25.13.5523-5534.2005
Esf2p, a U3-associated factor required for small-subunit processome assembly and compaction
Abstract
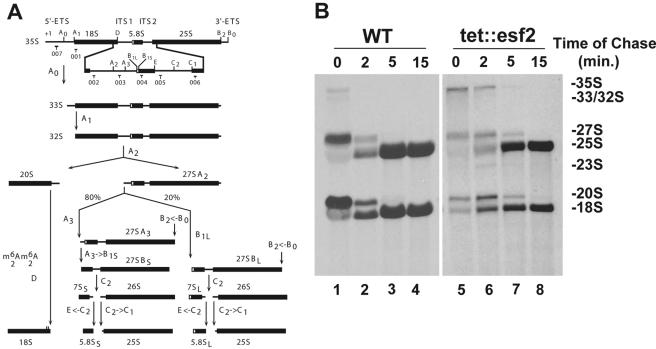
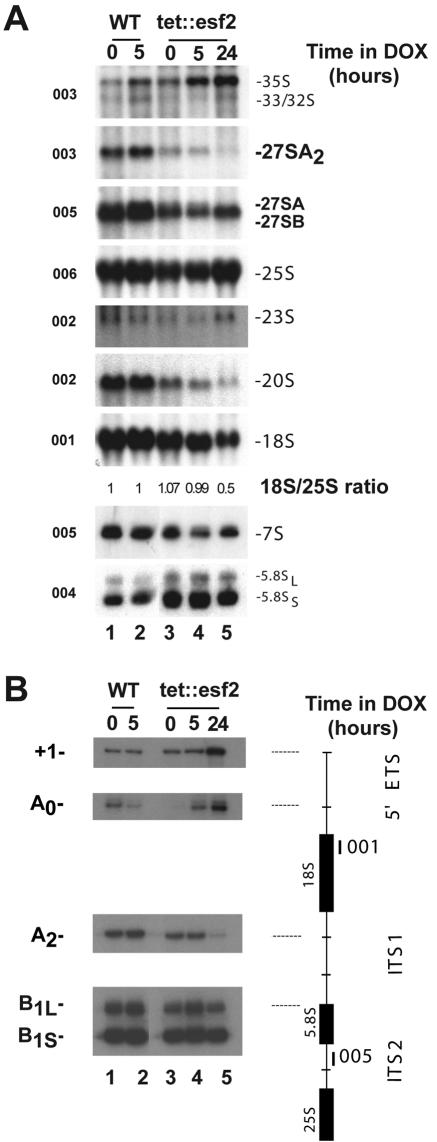
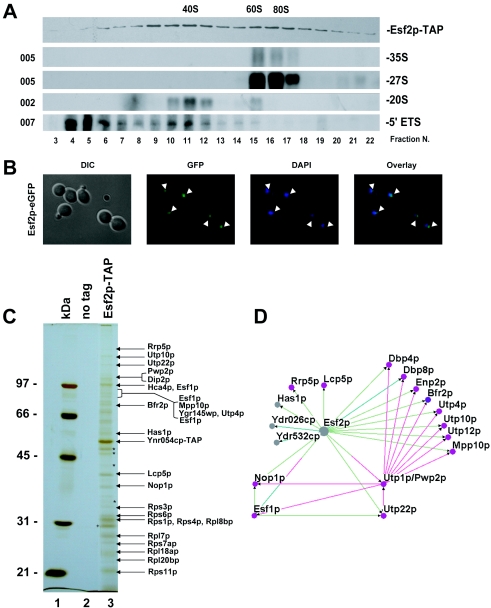
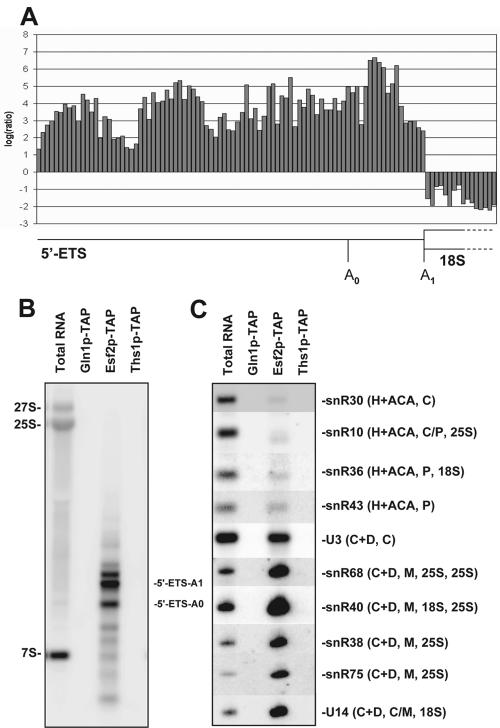
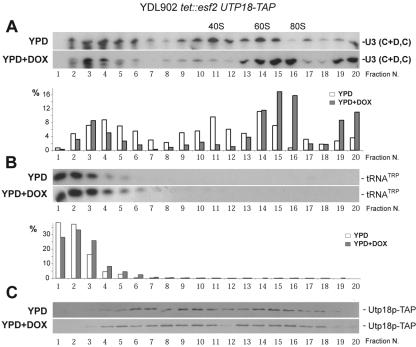
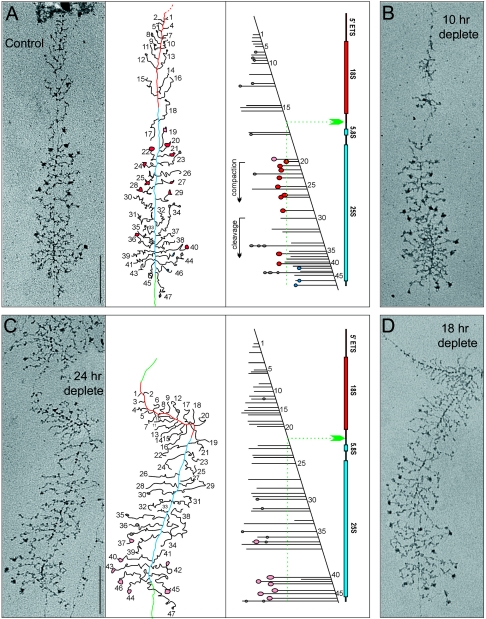
Esf2p is the Saccharomyces cerevisiae homolog of mouse ABT1, a protein previously identified as a putative partner of the TATA-element binding protein. However, large-scale studies have indicated that Esf2p is primarily localized to the nucleolus and that it physically associates with pre-rRNA processing factors. Here, we show that Esf2p-depleted cells are defective for pre-rRNA processing at the early nucleolar cleavage sites A0 through A2 and consequently are inhibited for 18S rRNA synthesis. Esf2p was stably associated with the 5' external transcribed spacer (ETS) and the box C+D snoRNA U3, as well as additional box C+D snoRNAs and proteins enriched within the small-subunit (SSU) processome/90S preribosomes. Esf2p colocalized on glycerol gradients with 90S preribosomes and slower migrating particles containing 5' ETS fragments. Strikingly, upon Esf2p depletion, chromatin spreads revealed that SSU processome assembly and compaction are inhibited and glycerol gradient analysis showed that U3 remains associated within 90S preribosomes. This suggests that in the absence of proper SSU processome assembly, early pre-rRNA processing is inhibited and U3 is not properly released from the 35S pre-rRNAs. The identification of ABT1 in a large-scale analysis of the human nucleolar proteome indicates that its role may also be conserved in mammals.
Figures
References
-
- Andersen, J. S., C. E. Lyon, A. H. Fox, A. K. Leung, Y. W. Lam, H. Steen, M. Mann, and A. I. Lamond. 2002. Directed proteomic analysis of the human nucleolus. Curr. Biol. 12:1-11. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases