

The selective recruitment of mRNA to the ER and an increase in initiation are important for glucose-stimulated proinsulin synthesis in pancreatic beta-cells
- PMID: 15972000
- PMCID: PMC1276927
- DOI: 10.1042/BJ20050468
The selective recruitment of mRNA to the ER and an increase in initiation are important for glucose-stimulated proinsulin synthesis in pancreatic beta-cells
Abstract
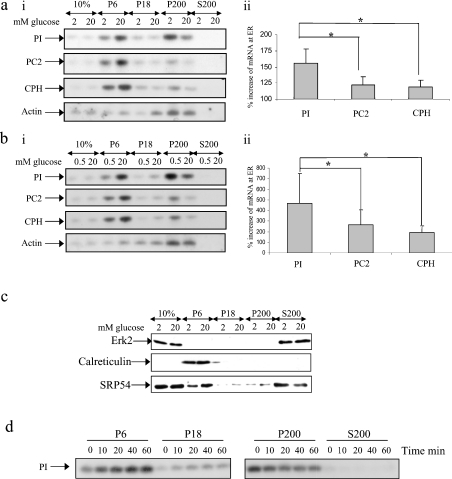
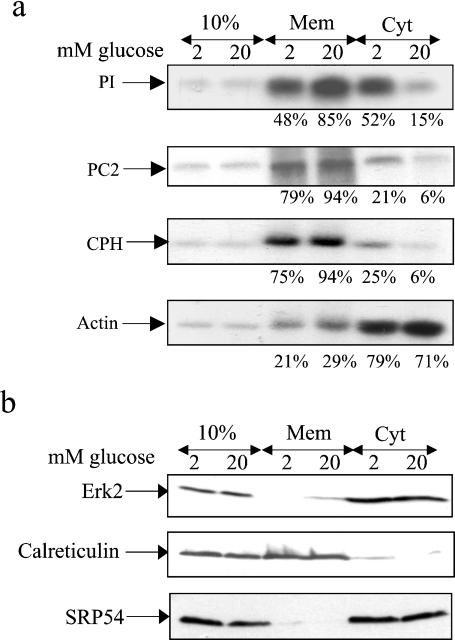
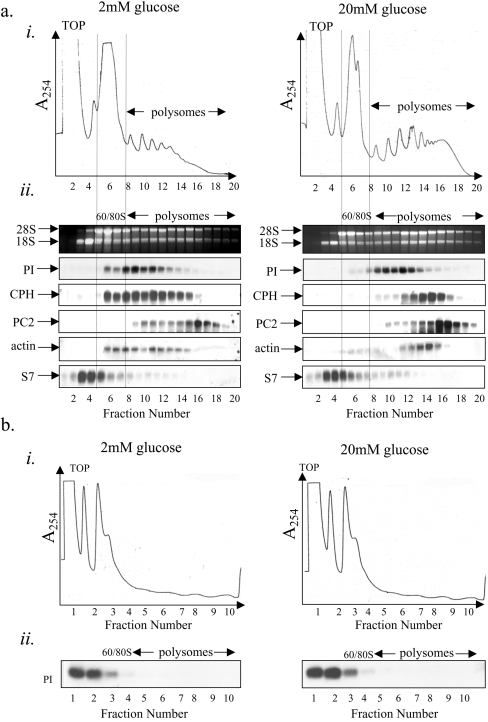
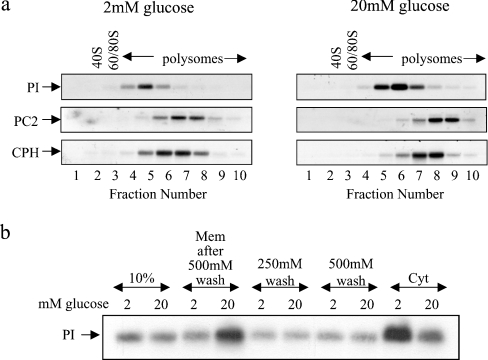
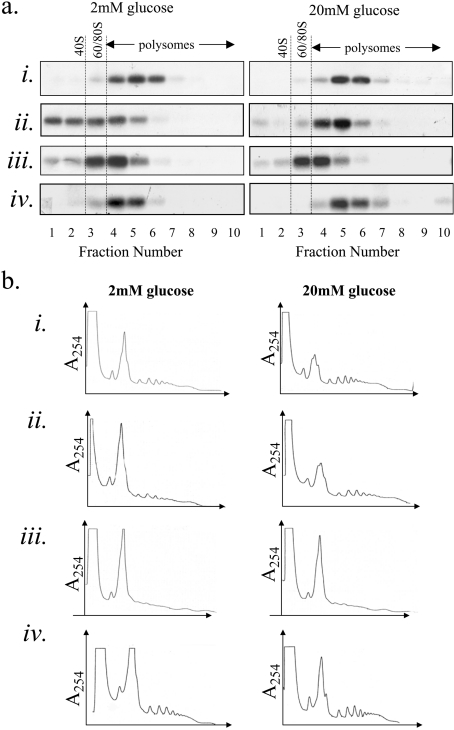
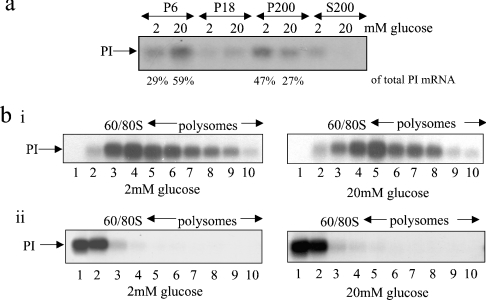
Glucose acutely stimulates proinsulin synthesis in pancreatic beta-cells through a poorly understood post-transcriptional mechanism. In the present study, we demonstrate in pancreatic beta-cells that glucose stimulates the recruitment of ribosome-associated proinsulin mRNA, located in the cytoplasm, to the ER (endoplasmic reticulum), the site of proinsulin synthesis, and that this plays an important role in glucose-stimulated proinsulin synthesis. Interestingly, glucose has greater stimulatory effect on the recruitment of proinsulin mRNA to the ER compared with other mRNAs encoding secretory proteins. This, as far as we are aware, is the first example whereby mRNAs encoding secretory proteins are selectively recruited to the ER and provides a novel regulatory mechanism for secretory protein synthesis. Contrary to previous reports, and importantly in understanding the mechanism by which glucose stimulates proinsulin synthesis, we demonstrate that there is no large pool of 'free' proinsulin mRNA in the cytoplasm and that glucose does not increase the rate of de novo initiation on the proinsulin mRNA. However, we show that glucose does stimulate the rate of ribosome recruitment on to ribosome-associated proinsulin mRNA. In conclusion, our results provide evidence that the selective recruitment of proinsulin mRNA to the ER, together with increases in the rate of initiation are important mediators of glucose-stimulated proinsulin synthesis in pancreatic beta-cells.
Figures

References
-
- Campbell I. L., Hellquist L. N., Taylor K. W. Insulin biosynthesis and its regulation. Clin. Sci. 1982;62:449–455. - PubMed
-
- Itoh N., Sei T., Nose K., Okamoto H. Glucose stimulation of the proinsulin synthesis in isolated pancreatic islets without increasing amount of proinsulin mRNA. FEBS Lett. 1978;93:343–347. - PubMed
-
- Itoh N., Okamoto H. Translational control of proinsulin synthesis by glucose. Nature (London) 1980;283:100–102. - PubMed
-
- Permutt M. A., Kipnis D. M. Insulin biosynthesis. I. On the mechanism of glucose stimulation. J. Biol. Chem. 1972;247:1194–1199. - PubMed
-
- Jahr H., Schroder D., Ziegler B., Ziegler M., Zuhlke H. Transcriptional and translational control of glucose-stimulated (pro)insulin biosynthesis. Eur. J. Biochem. 1980;110:499–505. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources