

A role for the Saccharomyces cerevisiae regulation of Ace2 and polarized morphogenesis signaling network in cell integrity
- PMID: 15972461
- PMCID: PMC1456762
- DOI: 10.1534/genetics.105.042101
A role for the Saccharomyces cerevisiae regulation of Ace2 and polarized morphogenesis signaling network in cell integrity
Abstract
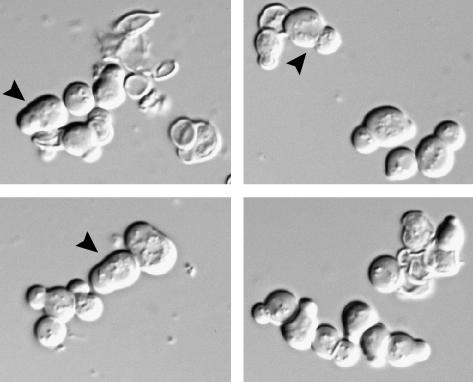
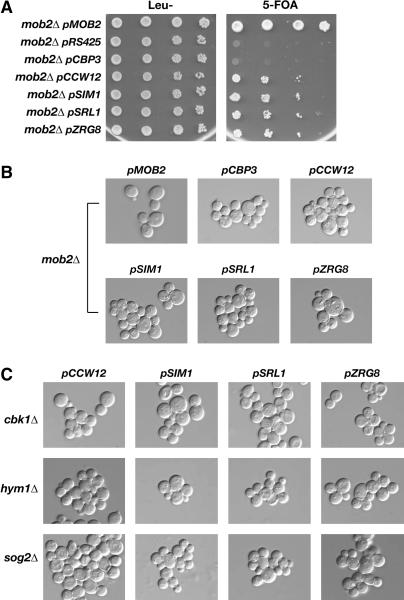
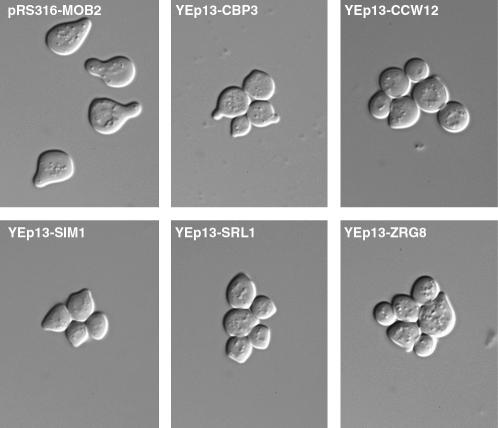
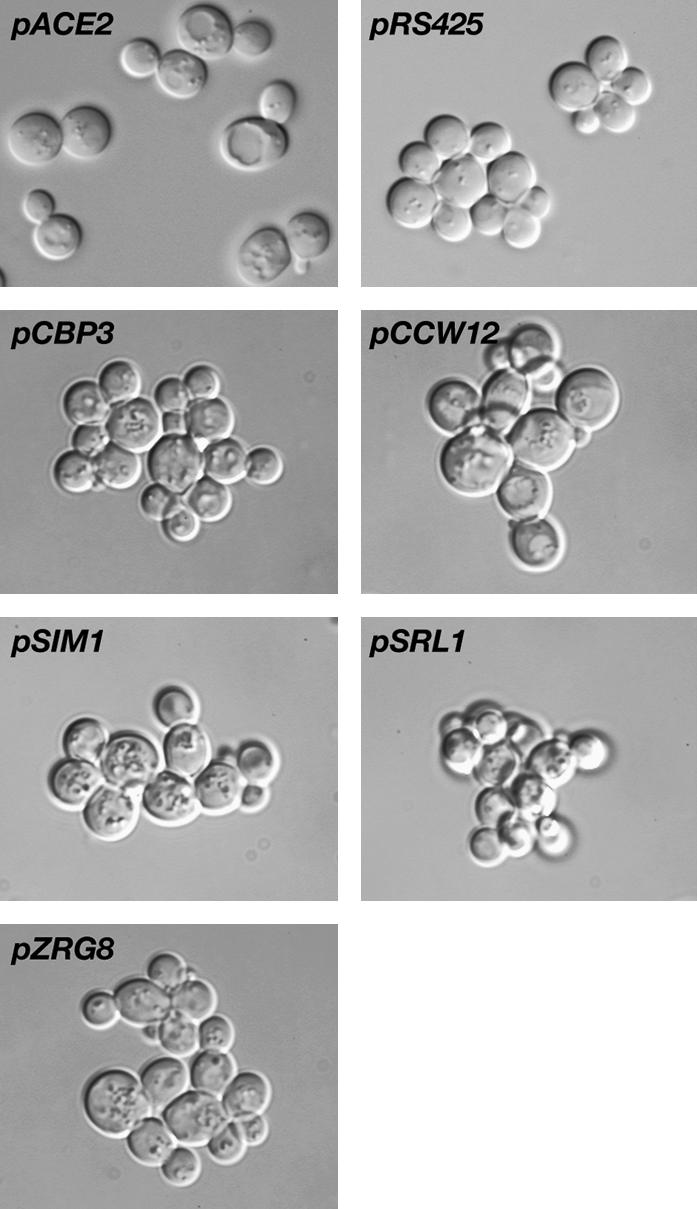
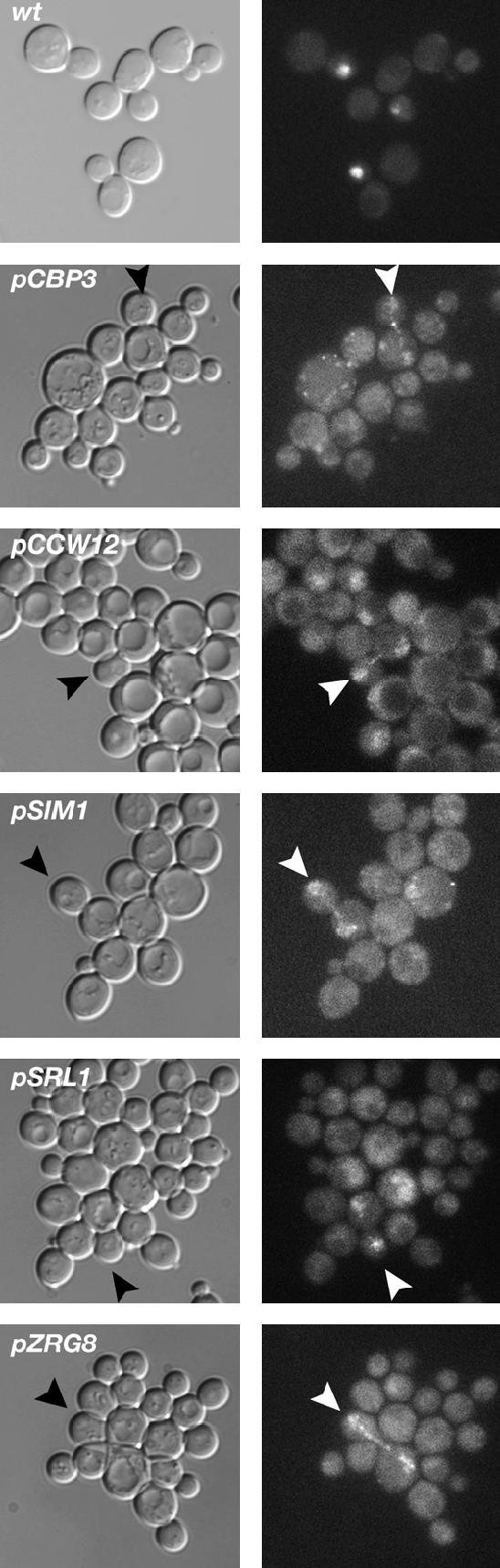
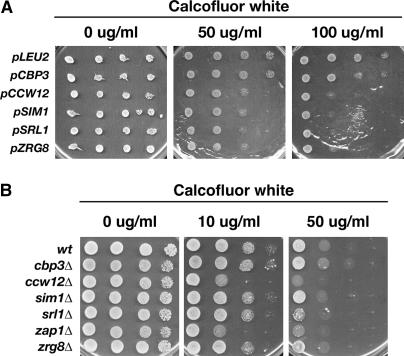
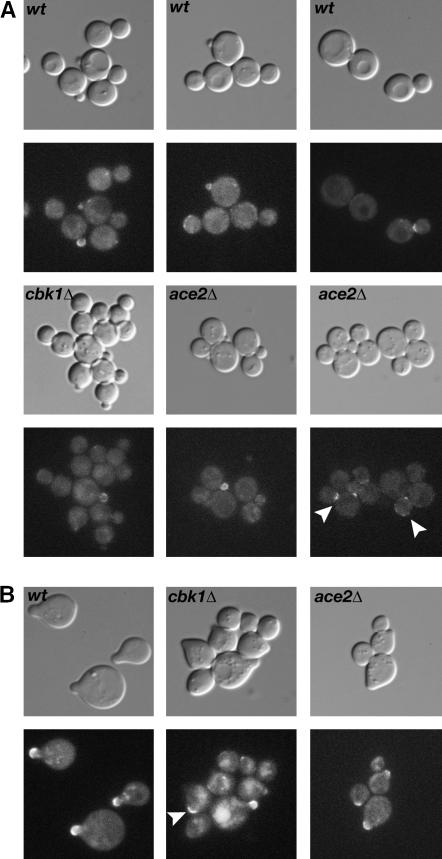
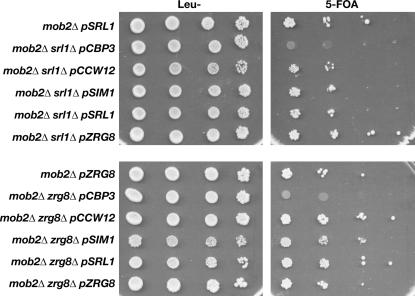
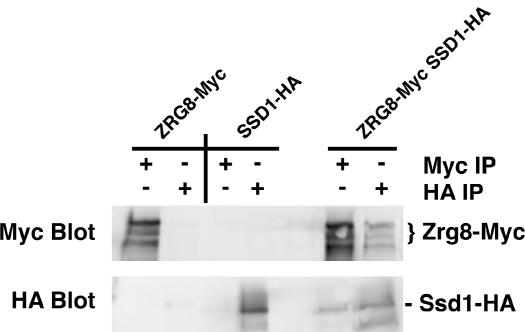
Saccharomyces cerevisiae RAM is a conserved signaling network that regulates maintenance of polarized growth and daughter-cell-specific transcription, the latter of which is critical for septum degradation. Consequently, cells defective in RAM function (designated ramDelta) are round in morphology, form feeble mating projections, and fail to separate following cytokinesis. It was recently demonstrated that RAM genes are essential in strains containing functional SSD1 (SSD1-v), which encodes a protein of unknown function that binds the RAM Cbk1p kinase. Here we investigated the essential function of RAM in SSD1-v strains and identified two functional groups of dosage suppressors for ramDelta lethality. We establish that all ramDelta mutants exhibit cell integrity defects and cell lysis. All dosage suppressors rescue the lysis but not the cell polarity or cell separation defects of ramDelta cells. One class of dosage suppressors is composed of genes encoding cell wall proteins, indicating that alterations in cell wall structure can rescue the cell lysis in ramDelta cells. Another class of ramDelta dosage suppressors is composed of ZRG8 and SRL1, which encode two unrelated proteins of unknown function. We establish that ZRG8 and SRL1 share similar genetic interactions and phenotypes. Significantly, Zrg8p coprecipitates with Ssd1p, localizes similarly to RAM proteins, and is dependent on RAM for localization. Collectively, these data indicate that RAM and Ssd1p function cooperatively to control cell integrity and suggest that Zrg8p and Srl1p function as nonessential inhibitors of Ssd1p.
Figures

Similar articles
-
The RAM network in pathogenic fungi.Eukaryot Cell. 2012 Jun;11(6):708-17. doi: 10.1128/EC.00044-12. Epub 2012 Apr 27. Eukaryot Cell. 2012. PMID: 22544903 Free PMC article. Review.
-
RAM: a conserved signaling network that regulates Ace2p transcriptional activity and polarized morphogenesis.Mol Biol Cell. 2003 Sep;14(9):3782-803. doi: 10.1091/mbc.e03-01-0018. Epub 2003 May 29. Mol Biol Cell. 2003. PMID: 12972564 Free PMC article.
-
The Saccharomyces cerevisiae Mob2p-Cbk1p kinase complex promotes polarized growth and acts with the mitotic exit network to facilitate daughter cell-specific localization of Ace2p transcription factor.J Cell Biol. 2002 Sep 2;158(5):885-900. doi: 10.1083/jcb.200203094. Epub 2002 Aug 26. J Cell Biol. 2002. PMID: 12196508 Free PMC article.
-
Mutations in the Saccharomyces cerevisiae kinase Cbk1p lead to a fertility defect that can be suppressed by the absence of Brr1p or Mpt5p (Puf5p), proteins involved in RNA metabolism.Genetics. 2009 Sep;183(1):161-73. doi: 10.1534/genetics.109.105130. Epub 2009 Jun 22. Genetics. 2009. PMID: 19546315 Free PMC article.
-
Mitotic exit and separation of mother and daughter cells.Genetics. 2012 Dec;192(4):1165-202. doi: 10.1534/genetics.112.145516. Genetics. 2012. PMID: 23212898 Free PMC article. Review.
Cited by
-
The RAM network in pathogenic fungi.Eukaryot Cell. 2012 Jun;11(6):708-17. doi: 10.1128/EC.00044-12. Epub 2012 Apr 27. Eukaryot Cell. 2012. PMID: 22544903 Free PMC article. Review.
-
The yeast Cbk1 kinase regulates mRNA localization via the mRNA-binding protein Ssd1.J Cell Biol. 2011 Feb 21;192(4):583-98. doi: 10.1083/jcb.201011061. J Cell Biol. 2011. PMID: 21339329 Free PMC article.
-
Diverse RNA-binding proteins interact with functionally related sets of RNAs, suggesting an extensive regulatory system.PLoS Biol. 2008 Oct 28;6(10):e255. doi: 10.1371/journal.pbio.0060255. PLoS Biol. 2008. PMID: 18959479 Free PMC article.
-
Roles of the RAM signaling network in cell cycle progression in Saccharomyces cerevisiae.Curr Genet. 2006 Jun;49(6):384-92. doi: 10.1007/s00294-006-0069-y. Epub 2006 Mar 22. Curr Genet. 2006. PMID: 16552603
-
Identification of Morphogenesis-Related NDR Kinase Signaling Network and Its Regulation on Cold Tolerance in Maize.Plants (Basel). 2023 Oct 21;12(20):3639. doi: 10.3390/plants12203639. Plants (Basel). 2023. PMID: 37896102 Free PMC article.
References
-
- Bogomolnaya, L. M., R. Pathak, J. Guo, R. Cham, R. Aramayo et al., 2004. Hym1p affects cell cycle progression in Saccharomyces cerevisiae. Curr. Genet. 46: 183–192. - PubMed
-
- Breton, A. M., J. Schaeffer and M. Aigle, 2001. The yeast Rvs161 and Rvs167 proteins are involved in secretory vesicles targeting the plasma membrane and in cell integrity. Yeast 18: 1053–1068. - PubMed
-
- Colman-Lerner, A., T. E. Chin and R. Brent, 2001. Yeast Cbk1 and Mob2 activate daughter-specific genetic programs to induce asymmetric cell fates. Cell 107: 739–750. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
