

Novel innate immune functions for galectin-1: galectin-1 inhibits cell fusion by Nipah virus envelope glycoproteins and augments dendritic cell secretion of proinflammatory cytokines
- PMID: 15972675
- PMCID: PMC4428613
- DOI: 10.4049/jimmunol.175.1.413
Novel innate immune functions for galectin-1: galectin-1 inhibits cell fusion by Nipah virus envelope glycoproteins and augments dendritic cell secretion of proinflammatory cytokines
Abstract
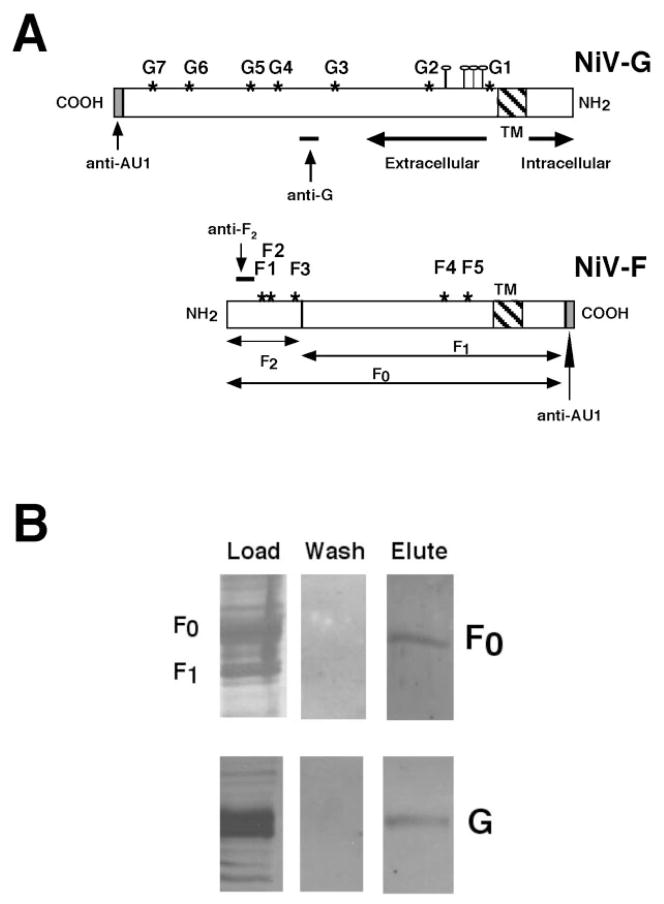
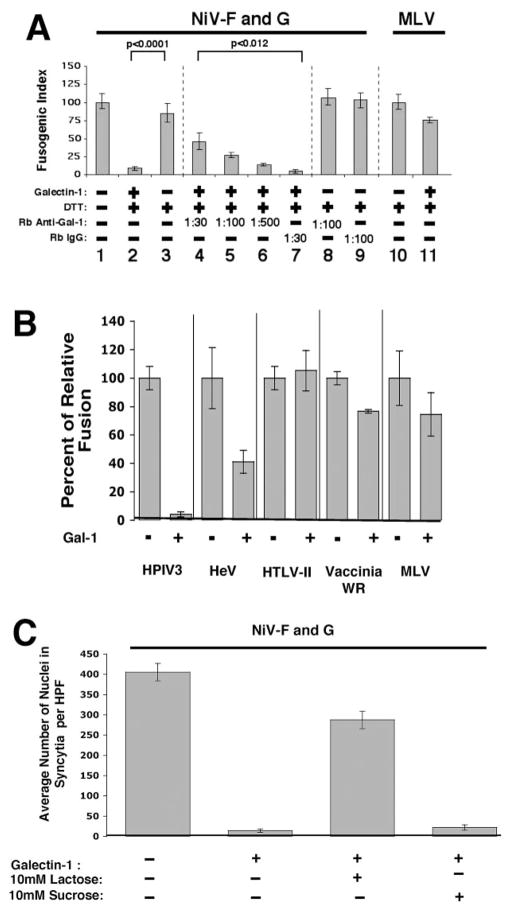
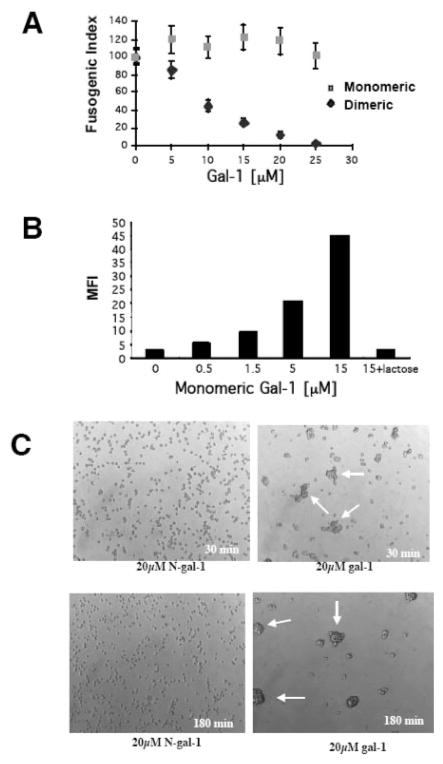
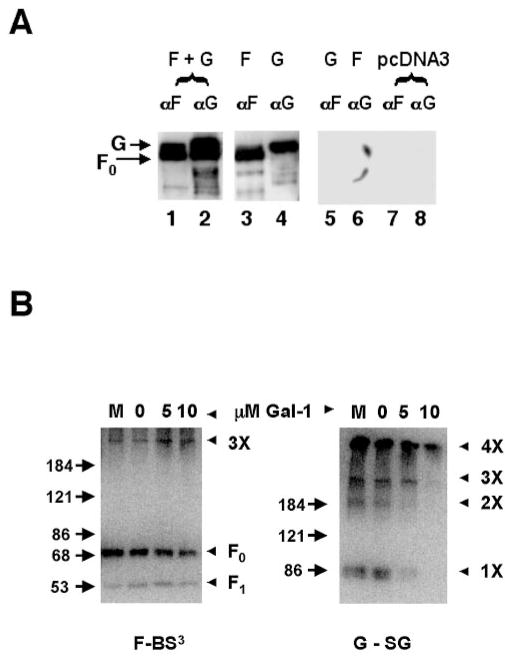
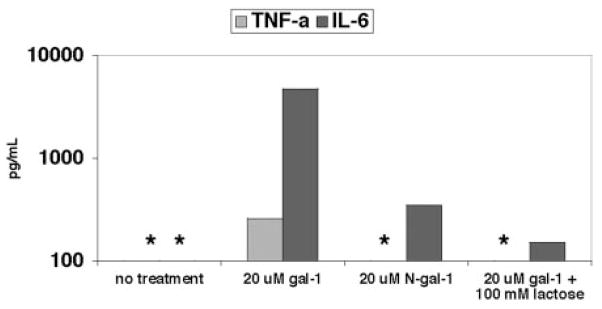
Galectin-1 (gal-1), an endogenous lectin secreted by a variety of cell types, has pleiotropic immunomodulatory functions, including regulation of lymphocyte survival and cytokine secretion in autoimmune, transplant disease, and parasitic infection models. However, the role of gal-1 in viral infections is unknown. Nipah virus (NiV) is an emerging pathogen that causes severe, often fatal, febrile encephalitis. The primary targets of NiV are endothelial cells. NiV infection of endothelial cells results in cell-cell fusion and syncytia formation triggered by the fusion (F) and attachment (G) envelope glycoproteins of NiV that bear glycan structures recognized by gal-1. In the present study, we report that NiV envelope-mediated cell-cell fusion is blocked by gal-1. This inhibition is specific to the Paramyxoviridae family because gal-1 did not inhibit fusion triggered by envelope glycoproteins of other viruses, including two retroviruses and a pox virus, but inhibited fusion triggered by envelope glycoproteins of the related Hendra virus and another paramyxovirus. The physiologic dimeric form of gal-1 is required for fusion inhibition because a monomeric gal-1 mutant had no inhibitory effect on cell fusion. gal-1 binds to specific N-glycans on NiV glycoproteins and aberrantly oligomerizes NiV-F and NiV-G, indicating a mechanism for fusion inhibition. gal-1 also increases dendritic cell production of proinflammatory cytokines such as IL-6, known to be protective in the setting of other viral diseases such as Ebola infections. Thus, gal-1 may have direct antiviral effects and may also augment the innate immune response against this emerging pathogen.
Conflict of interest statement
The authors have no financial conflict of interest.
Figures
Similar articles
-
Envelope-receptor interactions in Nipah virus pathobiology.Ann N Y Acad Sci. 2007 Apr;1102(1):51-65. doi: 10.1196/annals.1408.004. Ann N Y Acad Sci. 2007. PMID: 17470911 Free PMC article. Review.
-
Timing of galectin-1 exposure differentially modulates Nipah virus entry and syncytium formation in endothelial cells.J Virol. 2015 Mar;89(5):2520-9. doi: 10.1128/JVI.02435-14. Epub 2014 Dec 10. J Virol. 2015. PMID: 25505064 Free PMC article.
-
N-glycans on Nipah virus fusion protein protect against neutralization but reduce membrane fusion and viral entry.J Virol. 2006 May;80(10):4878-89. doi: 10.1128/JVI.80.10.4878-4889.2006. J Virol. 2006. PMID: 16641279 Free PMC article.
-
Novel Functions of Hendra Virus G N-Glycans and Comparisons to Nipah Virus.J Virol. 2015 Jul;89(14):7235-47. doi: 10.1128/JVI.00773-15. Epub 2015 May 6. J Virol. 2015. PMID: 25948743 Free PMC article.
-
Molecular characteristics of the Nipah virus glycoproteins.Ann N Y Acad Sci. 2007 Apr;1102:39-50. doi: 10.1196/annals.1408.003. Ann N Y Acad Sci. 2007. PMID: 17470910 Review.
Cited by
-
Antitumor agent calixarene 0118 targets human galectin-1 as an allosteric inhibitor of carbohydrate binding.J Med Chem. 2012 Jun 14;55(11):5121-9. doi: 10.1021/jm300014q. Epub 2012 May 30. J Med Chem. 2012. PMID: 22575017 Free PMC article.
-
Polybasic KKR motif in the cytoplasmic tail of Nipah virus fusion protein modulates membrane fusion by inside-out signaling.J Virol. 2007 May;81(9):4520-32. doi: 10.1128/JVI.02205-06. Epub 2007 Feb 14. J Virol. 2007. PMID: 17301148 Free PMC article.
-
Human galectin-1 and galectin-3 promote Tropheryma whipplei infection.Gut Microbes. 2021 Jan-Dec;13(1):1-15. doi: 10.1080/19490976.2021.1884515. Gut Microbes. 2021. PMID: 33573443 Free PMC article.
-
Functions of cell surface galectin-glycoprotein lattices.Curr Opin Struct Biol. 2007 Oct;17(5):513-20. doi: 10.1016/j.sbi.2007.09.002. Epub 2007 Oct 22. Curr Opin Struct Biol. 2007. PMID: 17950594 Free PMC article. Review.
-
Galectin-1 regulates tissue exit of specific dendritic cell populations.J Biol Chem. 2015 Sep 11;290(37):22662-77. doi: 10.1074/jbc.M115.644799. Epub 2015 Jul 27. J Biol Chem. 2015. PMID: 26216879 Free PMC article.
References
-
- Chua KB, Bellini WJ, Rota PA, Harcourt BH, Tamin A, Lam SK, Ksiazek TG, Rollin PE, Zaki SR, Shieh W, et al. Nipah virus: a recently emergent deadly paramyxovirus. Science. 2000;288:1432–1435. - PubMed
-
- Lam SK. Nipah virus: a potential agent of bioterrorism? Antiviral Res. 2003;57:113–119. - PubMed
-
- Nipah virus outbreak(s) in Bangladesh, January–April 2004. Wkly Epidemiol Rec. 2004;79:168. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
