

Alanine scanning of Arp1 delineates a putative binding site for Jnm1/dynamitin and Nip100/p150Glued
- PMID: 15975903
- PMCID: PMC1196314
- DOI: 10.1091/mbc.e05-02-0093
Alanine scanning of Arp1 delineates a putative binding site for Jnm1/dynamitin and Nip100/p150Glued
Abstract
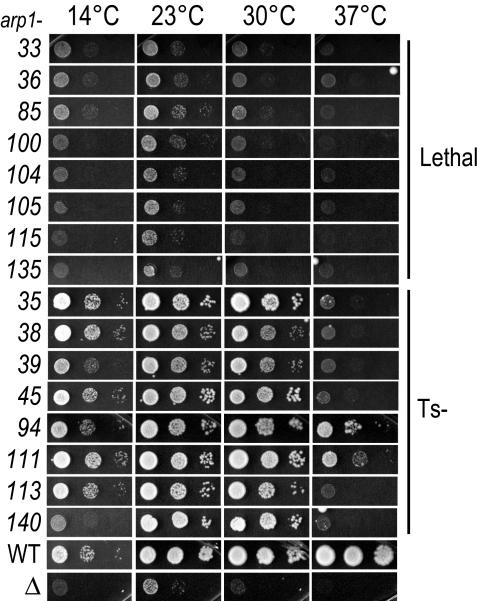
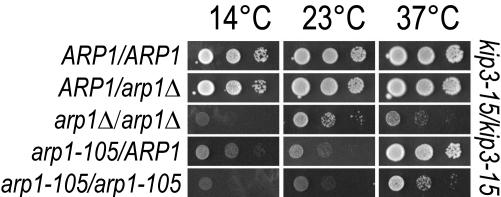
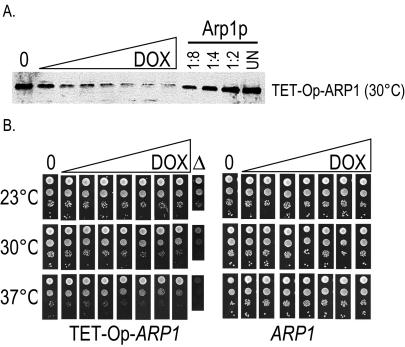
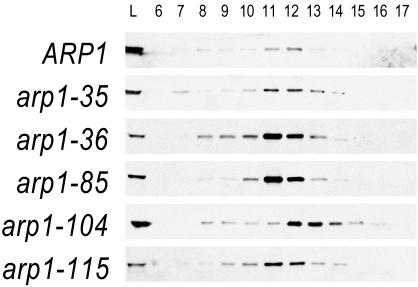
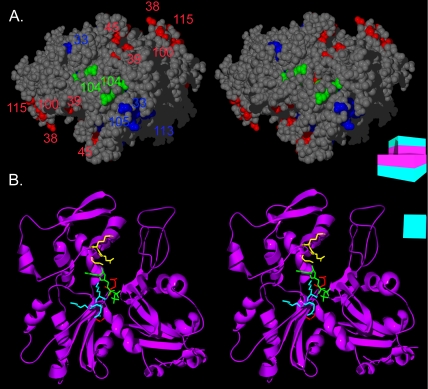
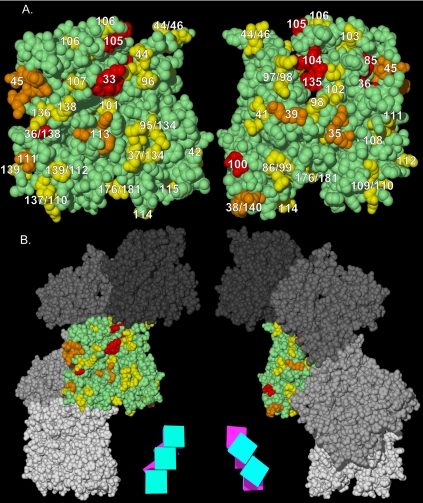
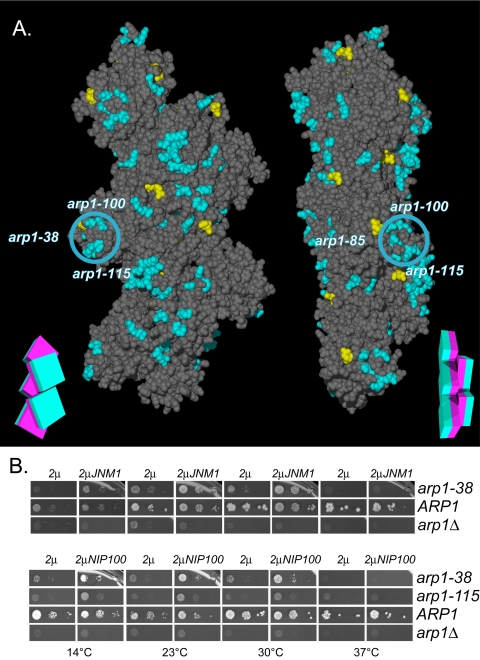
Arp1p is the only actin-related protein (ARP) known to form actin-like filaments. Unlike actin, Arp1p functions with microtubules, as part of the dynein regulator, dynactin. Arp1p's dissimilar functions imply interactions with a distinct set of proteins. To distinguish surface features relating to Arp1p's core functions and to identify the footprint of protein interactions essential for dynactin function, we performed the first complete charge-cluster-to-alanine scanning mutagenesis of an ARP and compared the results with a similar study of actin. The Arp1p mutations revealed three nonoverlapping surfaces with distinct genetic properties. One of these surfaces encompassed a region unique to Arp1p that is crucial for Jnm1p (dynamitin/p50) and Nip100p (p150(Glued)) association as well as pointed-end associations. Unlike the actin mutations, none of the ARP1 alleles disrupt filament formation; however, one pointed-end allele delayed the elution of Arp1p on gel filtration, consistent with loss of additional subunits.
Figures


References
-
- Amberg, D. C., Basart, E., and Botstein, D. (1995). Defining protein interactions with yeast actin in vivo. Nat. Struct. Biol. 2, 28–35. - PubMed
-
- Belli, G., Gari, E., Aldea, M., and Herrero, E. (1998a). Functional analysis of yeast essential genes using a promoter-substitution cassette and the tetracycline-regulatable dual expression system. Yeast 14, 1127–1138. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
