

The novel fission yeast protein Pal1p interacts with Hip1-related Sla2p/End4p and is involved in cellular morphogenesis
- PMID: 15975911
- PMCID: PMC1196324
- DOI: 10.1091/mbc.e04-11-0976
The novel fission yeast protein Pal1p interacts with Hip1-related Sla2p/End4p and is involved in cellular morphogenesis
Abstract
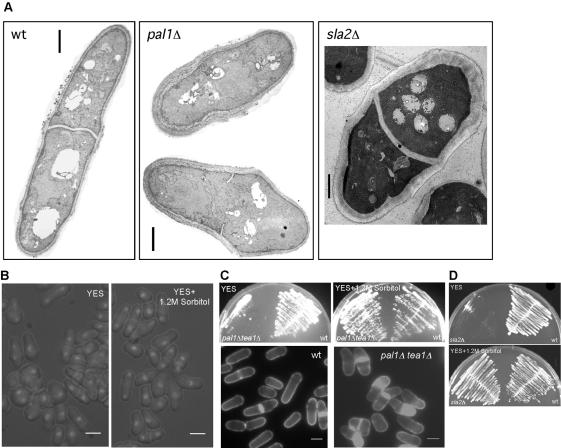
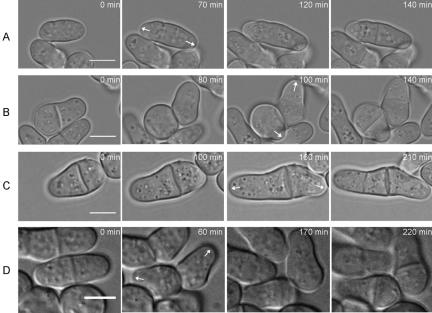
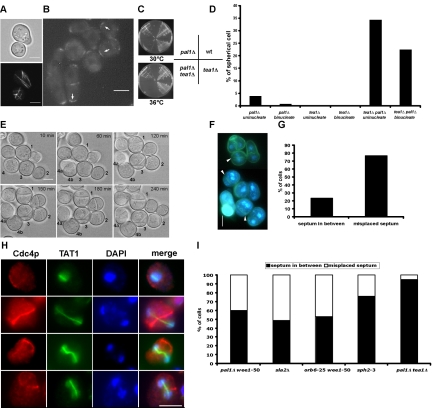
The establishment and maintenance of characteristic cellular morphologies is a fundamental property of all cells. Here we describe Schizosaccharomyces pombe Pal1p, a protein important for maintenance of cylindrical cellular morphology. Pal1p is a novel membrane-associated protein that localizes to the growing tips of interphase cells and to the division site in cells undergoing cytokinesis in an F-actin- and microtubule-independent manner. Cells deleted for pal1 display morphological defects, characterized by the occurrence of spherical and pear-shaped cells with an abnormal cell wall. Pal1p physically interacts and displays overlapping localization with the Huntingtin-interacting-protein (Hip1)-related protein Sla2p/End4p, which is also required for establishment of cylindrical cellular morphology. Sla2p is important for efficient localization of Pal1p to the sites of polarized growth and appears to function upstream of Pal1p. Interestingly, spherical pal1Delta mutants polarize to establish a pearlike morphology before mitosis in a manner dependent on the kelch-repeat protein Tea1p and the cell cycle inhibitory kinase Wee1p. Thus, overlapping mechanisms involving Pal1p, Tea1p, and Sla2p contribute to the establishment of cylindrical cellular morphology, which is important for proper spatial regulation of cytokinesis.
Figures






References
-
- Arellano, M., Niccoli, T., and Nurse, P. (2002). Tea3p is a cell end marker activating polarized growth in Schizosaccharomyces pombe. Curr. Biol. 12, 751-756. - PubMed
-
- Balasubramanian, M. K., Helfman, D. M., and Hemmingsen, S. M. (1992). A new tropomyosin essential for cytokinesis in the fission yeast S. pombe. Nature 360, 84-87. - PubMed
-
- Balasubramanian, M. K., McCollum, D., and Gould, K. L. (1997). Cytokinesis in fission yeast Schizosaccharomyces pombe. Methods Enzymol. 283, 494-506. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
