

Distinct contributions of hippocampal NMDA and AMPA receptors to encoding and retrieval of one-trial place memory
- PMID: 15976073
- PMCID: PMC6724786
- DOI: 10.1523/JNEUROSCI.0698-05.2005
Distinct contributions of hippocampal NMDA and AMPA receptors to encoding and retrieval of one-trial place memory
Abstract
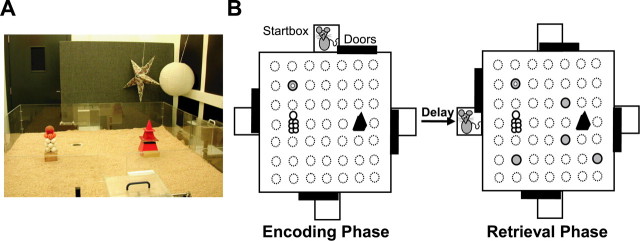
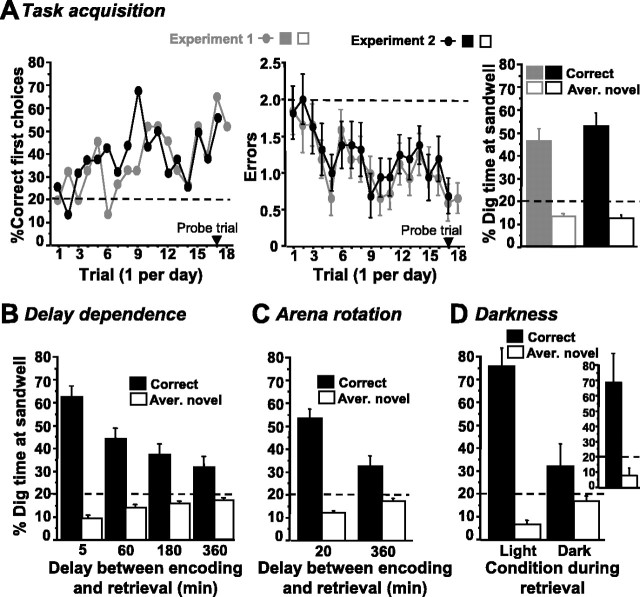
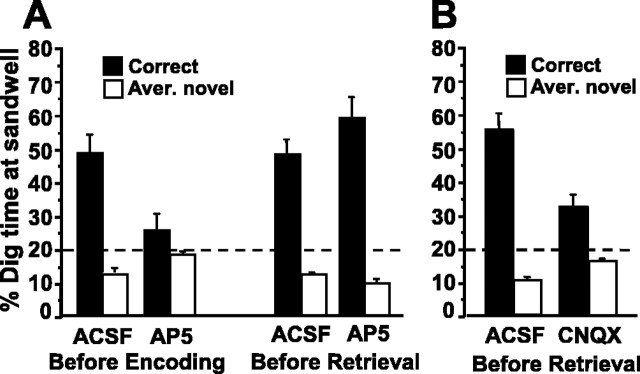
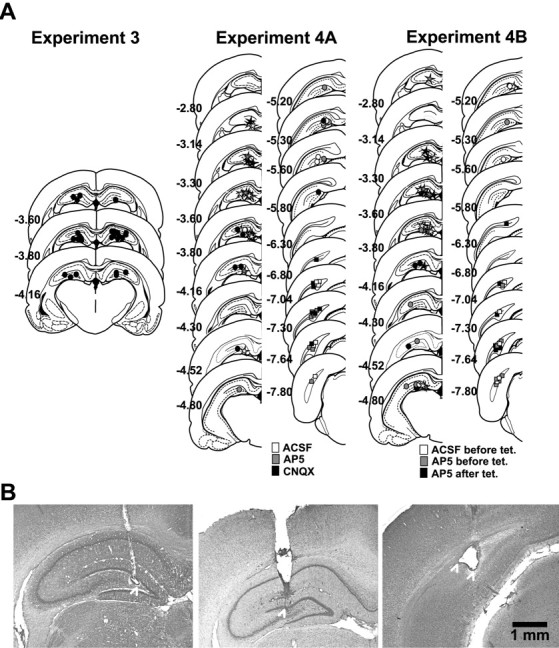
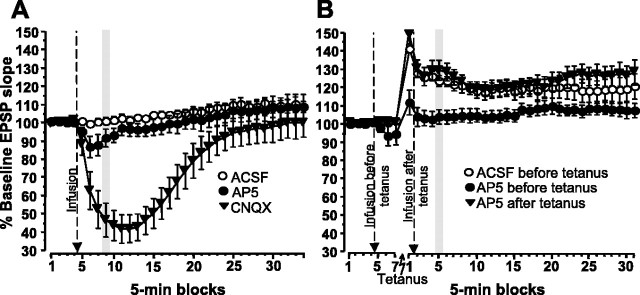
Allocentric place memory may serve to specify the context of events stored in human episodic memory. Recently, our laboratory demonstrated that, analogous to event-place associations in episodic memory, rats could associate, within one trial, a specific food flavor with an allocentrically defined place in an open arena. Encoding, but not retrieval, of such flavor-place associations required hippocampal NMDA receptors; retrieval depended on hippocampal AMPA receptors. This might have partly reflected the contributions of these receptors to encoding and retrieval of one-trial place, rather than flavor-place, memory. Therefore, the present study developed a food-reinforced arena paradigm to study encoding and retrieval of one-trial allocentric place memory in rats; memory relied on visuospatial information and declined with increasing retention delay, still being significant after 6 h, the longest delay tested (experiments 1 and 2). Hippocampal infusion of the NMDA receptor antagonist d-AP-5 blocked encoding without affecting retrieval; hippocampal infusion of the AMPA receptor antagonist CNQX impaired retrieval (experiment 3). Finally, we confirmed that the d-AP-5 infusions selectively blocked induction of long-term potentiation, a form of synaptic plasticity, whereas CNQX impaired fast excitatory transmission, at perforant-path dentate gyrus synapses in the dorsal hippocampus in vivo (experiment 4). Our results support that encoding, but not retrieval, of one-trial allocentric place memory requires the NMDA receptor-dependent induction of hippocampal synaptic plasticity, whereas retrieval depends on AMPA receptor-mediated fast excitatory hippocampal transmission. The contributions of hippocampal NMDA and AMPA receptors to one-trial allocentric place memory may be central to episodic memory and related episodic-like forms of memory in rats.
Figures
References
-
- Bannerman DM, Good MA, Butcher SP, Ramsay M, Morris RG (1995) Distinct components of spatial learning revealed by prior training and NMDA receptor blockade. Nature 378: 182-186. - PubMed
-
- Bannerman DM, Rawlins JNP, McHugh SB, Deacon RMJ, Yee BK, Bast T, Zhang W-N, Pothuizen HHJ, Feldon J (2004) Regional dissociations within the hippocampus-memory and anxiety. Neurosci Biobehav Rev 273-283. - PubMed
-
- Bast T, Feldon J (2003) Hippocampal modulation of sensorimotor processes. Prog Neurobiol 70: 319-345. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical