

Sexual dimorphism of short-wavelength photoreceptors in the small white butterfly, Pieris rapae crucivora
- PMID: 15976082
- PMCID: PMC6724796
- DOI: 10.1523/JNEUROSCI.1364-05.2005
Sexual dimorphism of short-wavelength photoreceptors in the small white butterfly, Pieris rapae crucivora
Abstract
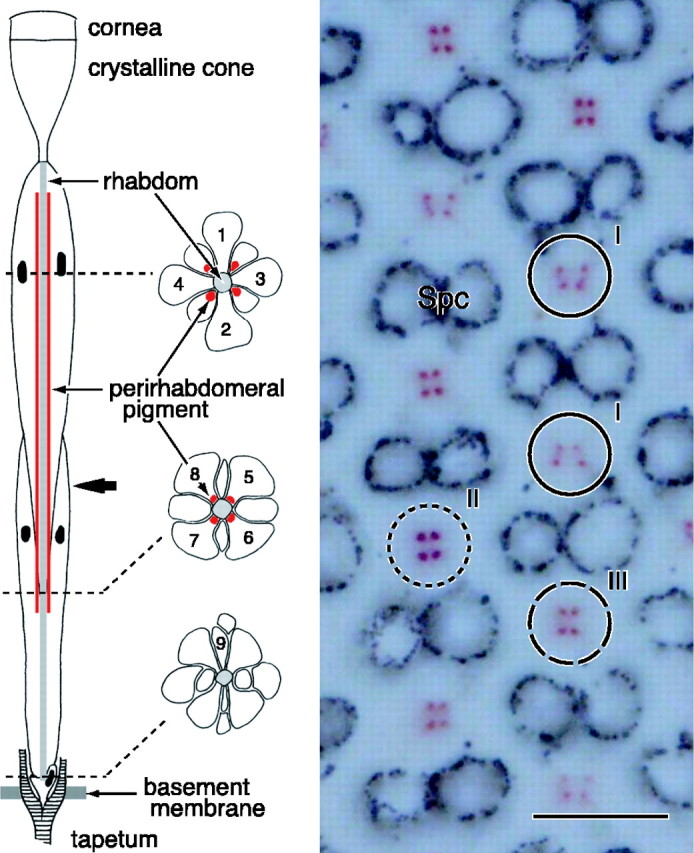
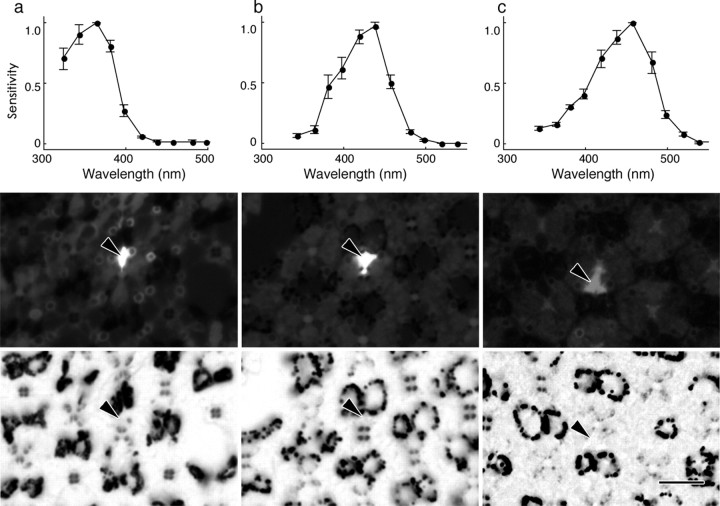
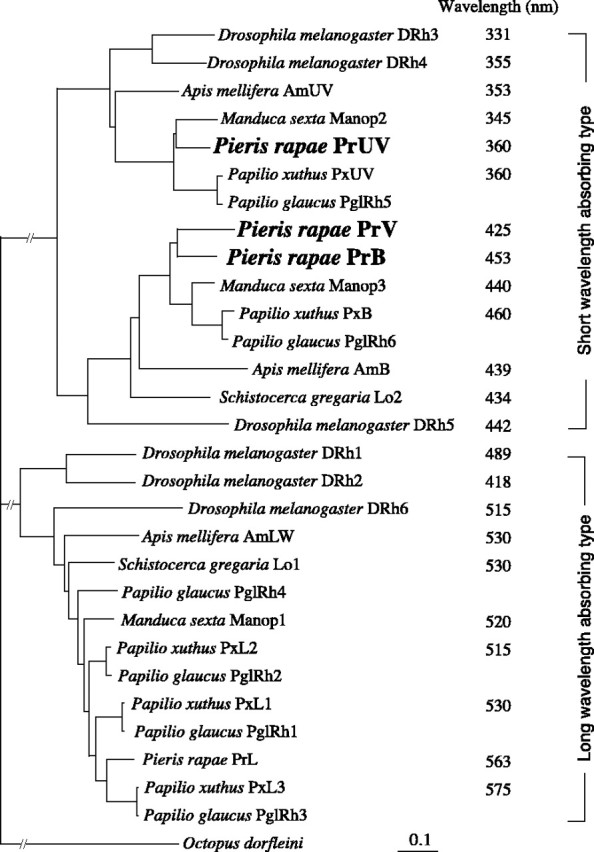
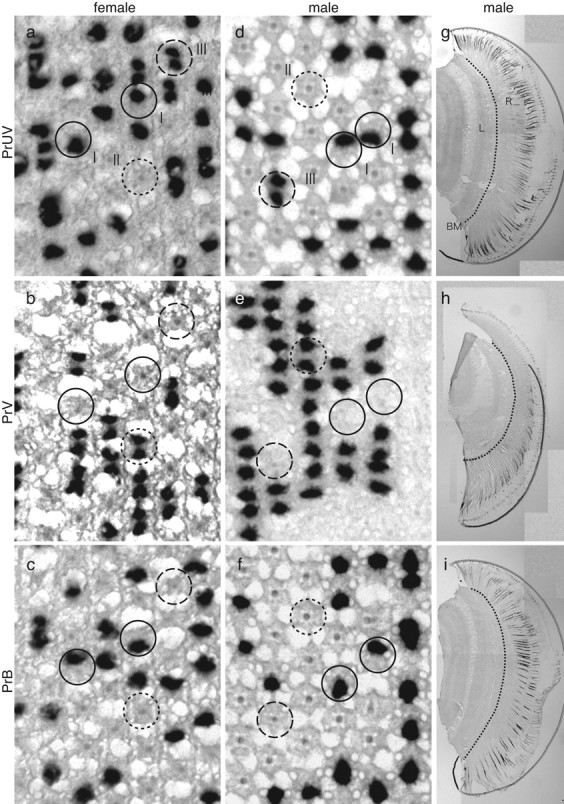
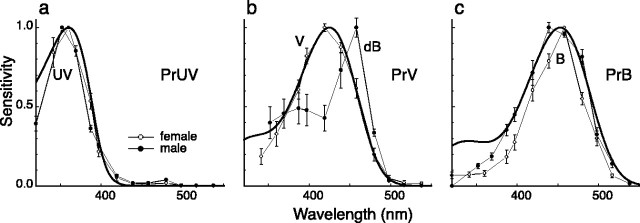
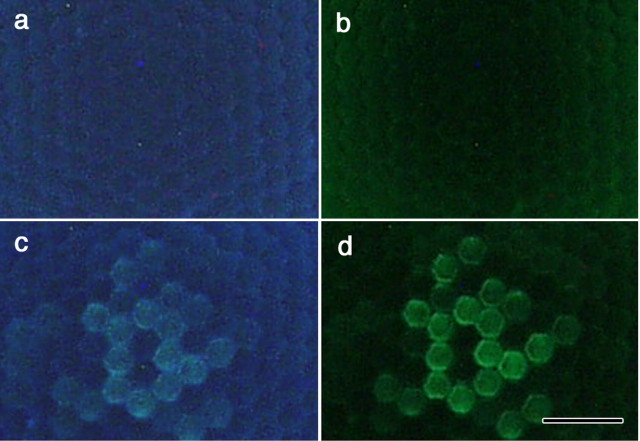
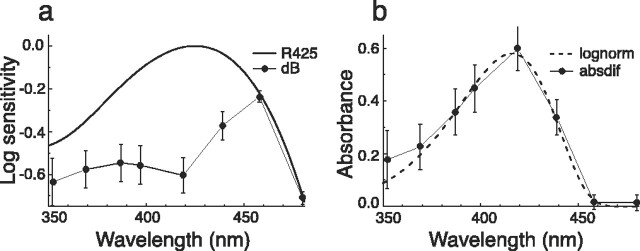
The eyes of the female small white butterfly, Pieris rapae crucivora, are furnished with three classes of short-wavelength photoreceptors, with sensitivity peaks in the ultraviolet (UV) (lambda(max) = 360 nm), violet (V) (lambda(max) = 425 nm), and blue (B) (lambda(max) = 453 nm) wavelength range. Analyzing the spectral origin of the photoreceptors, we isolated three novel mRNAs encoding opsins corresponding to short-wavelength-absorbing visual pigments. We localized the opsin mRNAs in the retinal tissue and found that each of the short-wavelength-sensitive photoreceptor classes exclusively expresses one of the opsin mRNAs. We, accordingly, termed the visual pigments PrUV, PrV, and PrB, respectively. The eyes of the male small white butterfly also use three classes of short-wavelength photoreceptors that equally uniquely express PrUV, PrV, and PrB. However, whereas the spectral sensitivities of the male photoreceptors with PrUV and PrB closely correspond to those of the female, the male photoreceptor expressing PrV has a double-peaked blue (dB) spectral sensitivity, strongly deviating from the spectral sensitivity of the female V photoreceptor. The male eyes contain a pigment that distinctly fluoresces under blue-violet as well as UV excitation light. It coexists with the dB photoreceptors and presumably acts as a spectral filter with an absorbance spectrum peaking at 416 nm. The narrow-band spectral sensitivity of the male dB photoreceptors probably evolved to improve the discrimination of the different wing colors of male and female P. rapae crucivora in the short-wavelength region of the spectrum.
Figures

References
-
- Arikawa K (2003) Spectral organization of the eye of a butterfly Papilio J Comp Physiol [A] 189: 791-800. - PubMed
-
- Arikawa K, Scholten DGW, Kinoshita M, Stavenga DG (1999a) Tuning of photoreceptor spectral sensitivities by red and yellow pigments in the butterfly Papilio xuthus Zool Sci 16: 17-24.
-
- Arikawa K, Mizuno S, Scholten DGW, Kinoshita M, Seki T, Kitamoto J, Stavenga DG (1999b) An ultraviolet absorbing pigment causes a narrow-band violet receptor and a single-peaked green receptor in the eye of the butterfly Papilio Vision Res 39: 1-8. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous