

Developmental changes in diffusion anisotropy coincide with immature oligodendrocyte progression and maturation of compound action potential
- PMID: 15976088
- PMCID: PMC6724805
- DOI: 10.1523/JNEUROSCI.4983-04.2005
Developmental changes in diffusion anisotropy coincide with immature oligodendrocyte progression and maturation of compound action potential
Abstract
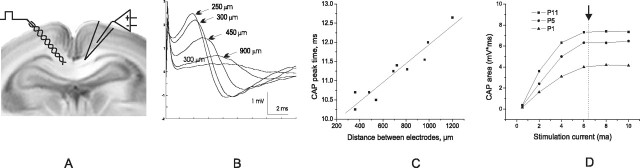
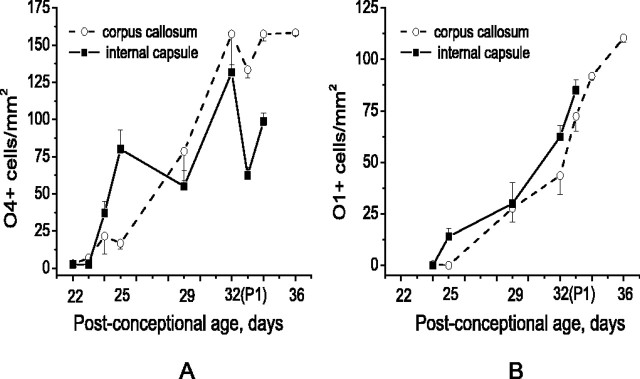
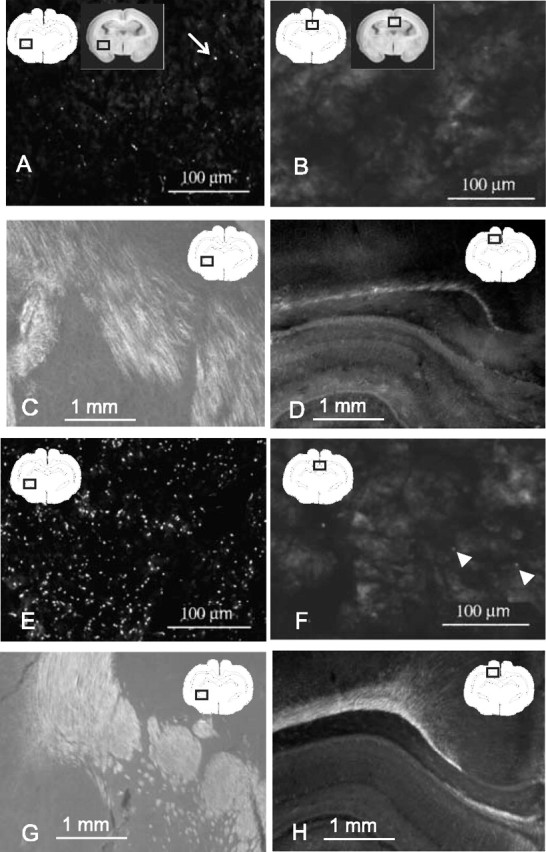
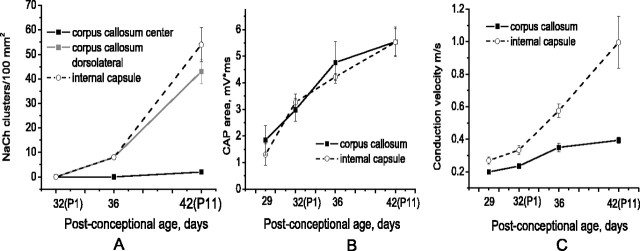
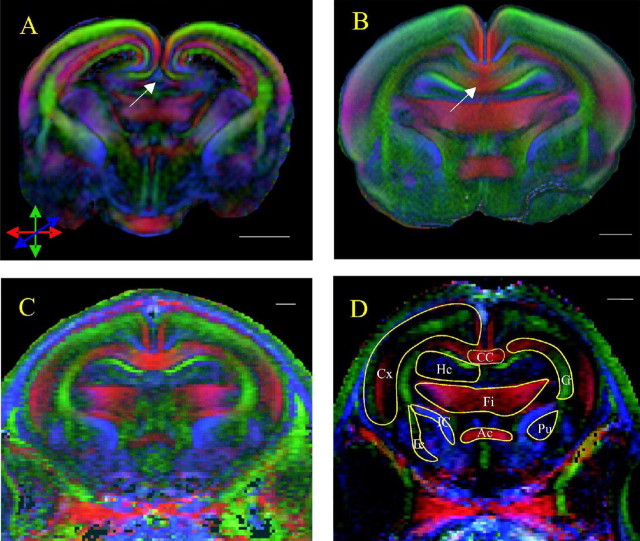
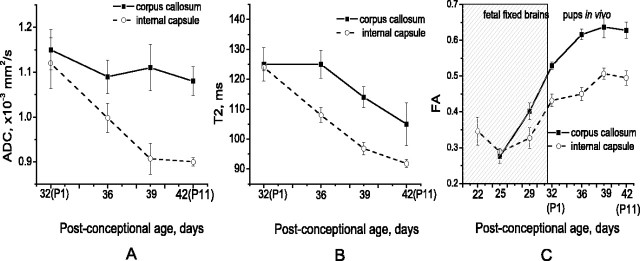
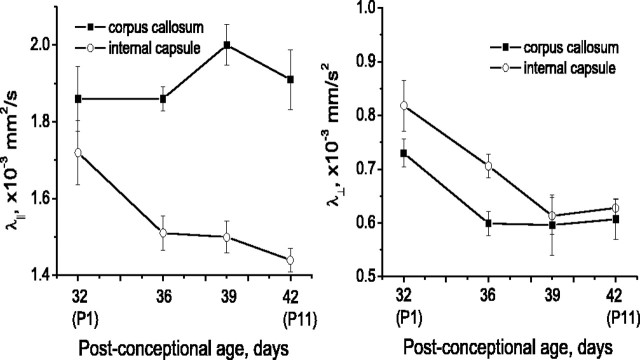
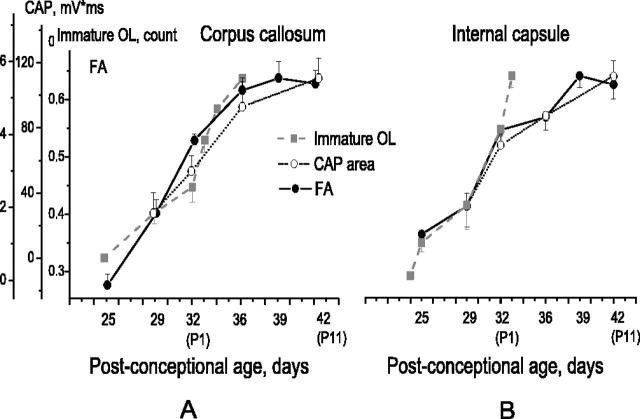
Disruption of oligodendrocyte lineage progression is implicated in the white-matter injury that occurs in cerebral palsy. We have previously published a model in rabbits consistent with cerebral palsy. Little is known of normal white-matter development in perinatal rabbits. Using a multidimensional approach, we defined the relationship of oligodendrocyte lineage progression and functional maturation of axons to structural development of selected cerebral white-matter tracts as determined by diffusion tensor imaging (DTI). Immunohistochemical studies showed that late oligodendrocyte progenitors appear at gestational age 22 [embryonic day 22 (E22)], whereas immature oligodendrocytes appear at E25, and both increase rapidly with time (approximately 13 cells/mm2/d) until the onset of myelination. Myelination began at postnatal day 5 (P5) (E36) in the internal capsule (IC) and at P11 in the medial corpus callosum (CC), as determined by localization of sodium channels and myelin basic protein. DTI of the CC and IC showed that fractional anisotropy (FA) increased rapidly between E25 and P1 (E32) (11% per day) and plateaued (<5% per day) after the onset of myelination. Postnatal maturation of the compound action potential (CAP) showed a developmental pattern similar to FA, with a rapid rise between E29 and P5 (in the CC, 18% per day) and a slower rise from P5 to P11 (in the CC, <5% per day). The development of immature oligodendrocytes after E29 coincides with changes in FA and CAP area in both the CC and IC. These findings suggest that developmental expansion of immature oligodendrocytes during the premyelination period may be important in defining structural and functional maturation of the white matter.
Figures
References
-
- Back SA, Rivkees SA (2004) Emerging concepts in periventricular white matter injury. Semin Perinatol 28: 405-414. - PubMed
-
- Baker AJ, Phan N, Moulton RJ, Fehlings MG, Yucel Y, Zhao M, Liu E, Tian GF (2002) Attenuation of the electrophysiological function of the corpus callosum after fluid percussion injury in the rat. J Neurotrauma 19: 587-599. - PubMed
-
- Baratti C, Barnett AS, Pierpaoli C (1999) Comparative MR imaging study of brain maturation in kittens with T1, T2, and the trace of the diffusion tensor. Radiology 210: 133-142. - PubMed
-
- Basser PJ, Jones DK (2002) Diffusion-tensor MRI: theory, experimental design and data analysis—a technical review. NMR Biomed 15: 456-467. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous