

Vitamin B1 functions as an activator of plant disease resistance
- PMID: 15980201
- PMCID: PMC1176421
- DOI: 10.1104/pp.104.058693
Vitamin B1 functions as an activator of plant disease resistance
Abstract
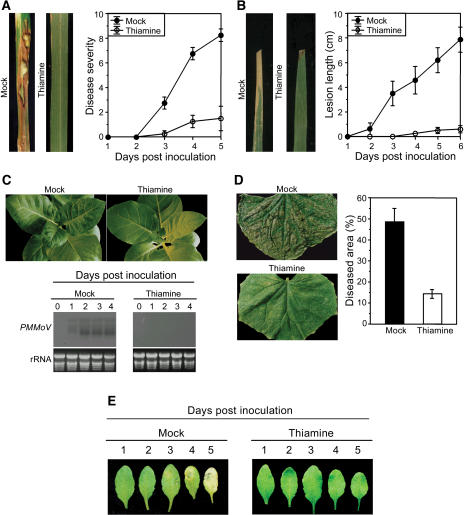
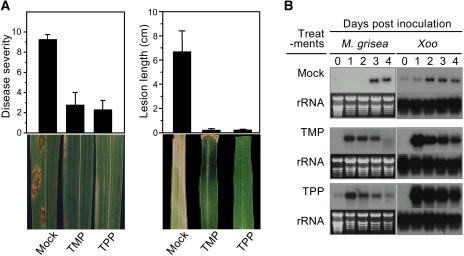
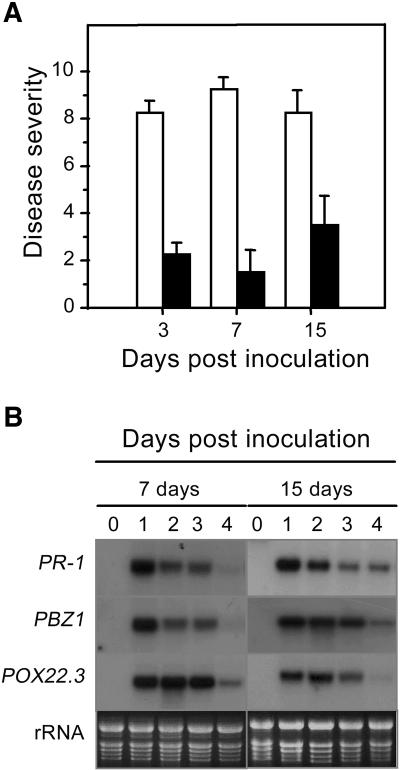
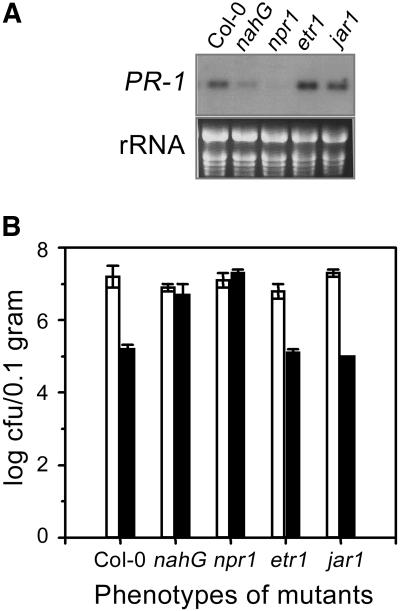
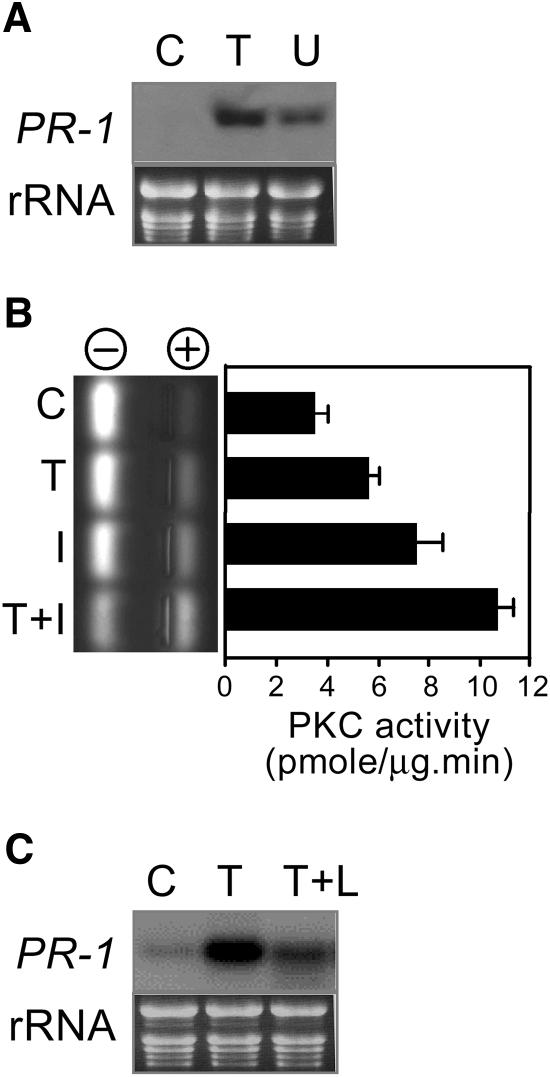
Vitamin B(1) (thiamine) is an essential nutrient for humans. Vitamin B(1) deficiency causes beriberi, which disturbs the central nervous and circulatory systems. In countries in which rice (Oryza sativa) is a major food, thiamine deficiency is prevalent because polishing of rice removes most of the thiamine in the grain. We demonstrate here that thiamine, in addition to its nutritional value, induces systemic acquired resistance (SAR) in plants. Thiamine-treated rice, Arabidopsis (Arabidopsis thaliana), and vegetable crop plants showed resistance to fungal, bacterial, and viral infections. Thiamine treatment induces the transient expression of pathogenesis-related (PR) genes in rice and other plants. In addition, thiamine treatment potentiates stronger and more rapid PR gene expression and the up-regulation of protein kinase C activity. The effects of thiamine on disease resistance and defense-related gene expression mobilize systemically throughout the plant and last for more than 15 d after treatment. Treatment of Arabidopsis ecotype Columbia-0 plants with thiamine resulted in the activation of PR-1 but not PDF1.2. Furthermore, thiamine prevented bacterial infection in Arabidopsis mutants insensitive to jasmonic acid or ethylene but not in mutants impaired in the SAR transduction pathway. These results clearly demonstrate that thiamine induces SAR in plants through the salicylic acid and Ca(2+)-related signaling pathways. The findings provide a novel paradigm for developing alternative strategies for the control of plant diseases.
Figures

References
-
- Achuo EA, Audenaert K, Meziane H, Hofte M (2004) The salicylic acid-dependent defence pathway is effective against different pathogens in tomato and tobacco. Plant Pathol 53: 65–72 - PubMed
-
- Ahn I-P, Park K, Kim C-H (2002) Rhizobacteria-induced resistance perturbs viral disease progress and triggers defense-related gene expression. Mol Cells 13: 302–308 - PubMed
-
- Allen RL, Bittner-Eddy PD, Grenville-Briggs LJ, Meitz JC, Rehmany AP, Rose LE, Beynon JL (2004) Host-parasite coevolutionary conflict between Arabidopsis and downy mildew. Science 306: 1957–1960 - PubMed
-
- Belkhadir Y, Nimchuk Z, Hubert DA, Mackey D, Dangl JL (2004) Arabidopsis RIN4 negatively regulates disease resistance mediated by RPS2 and RPM1 downstream or independent of the NDR1 signal modulator and is not required for the virulence functions of bacterial type III effectors AvrRpt2 or AvrRpm1. Plant Cell 16: 2822–2835 - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
