

Thermodynamic battle for photosynthate acquisition between sieve tubes and adjoining parenchyma in transport phloem
- PMID: 15980202
- PMCID: PMC1176423
- DOI: 10.1104/pp.104.058511
Thermodynamic battle for photosynthate acquisition between sieve tubes and adjoining parenchyma in transport phloem
Abstract
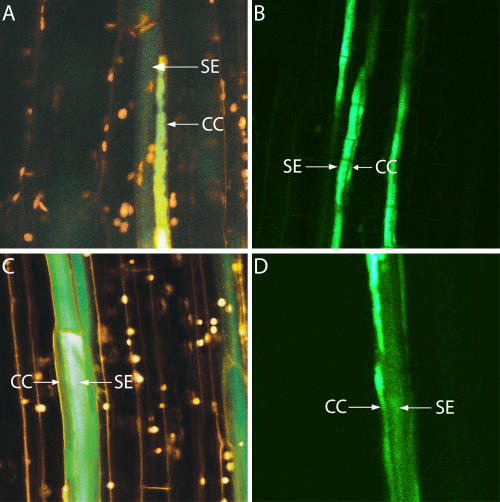
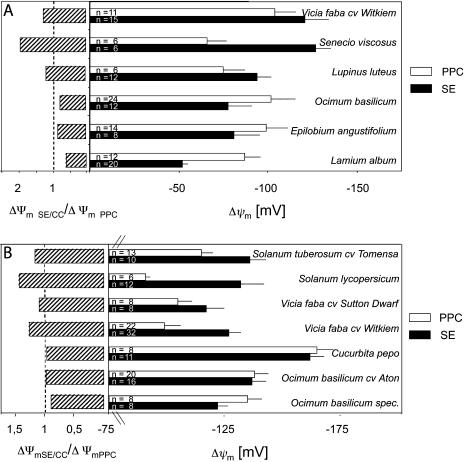
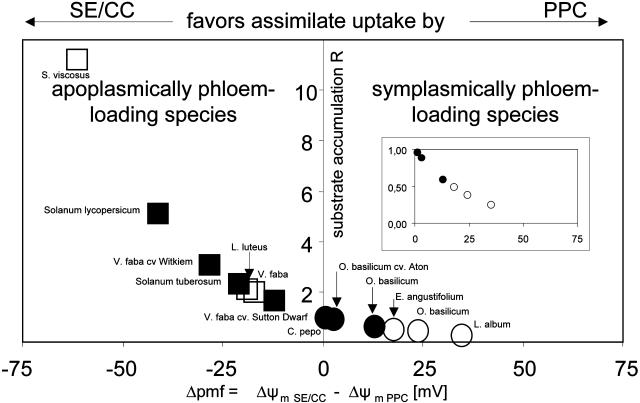
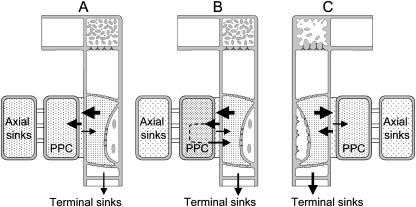
In transport phloem, photoassimilates escaping from the sieve tubes are released into the apoplasmic space between sieve element (SE)/companion cell (CC) complexes (SE/CCs) and phloem parenchyma cells (PPCs). For uptake respective retrieval, PPCs and SE/CCs make use of plasma membrane translocators energized by the proton motive force (PMF). Their mutual competitiveness, which essentially determines the amount of photoassimilates translocated through the sieve tubes, therefore depends on the respective PMFs. We measured the components of the PMF, membrane potential and DeltapH, of SE/CCs and PPCs in transport phloem. Membrane potentials of SE/CCs and PPCs in tissue slices as well as in intact plants fell into two categories. In the first group including apoplasmically phloem-loading species (e.g. Vicia, Solanum), the membrane potentials of the SEs are more negative than those of the PPCs. In the second group including symplasmically phloem-loading species (e.g. Cucurbita, Ocimum), membrane potentials of SEs are equal to or slightly more positive than those of PPCs. Pure sieve tube sap collected from cut aphid stylets was measured with H(+)-selective microelectrodes. Under our experimental conditions, pH of the sieve tube saps was around 7.5, which is comparable to the pH of cytoplasmic compartments in parenchymatous cells. In conclusion, only the membrane potential appears to be relevant for the PMF-determined competition between SE/CCs and PPCs. The findings may imply that the axial sinks along the pathway withdraw more photoassimilates from the sieve tubes in symplasmically loading species than in apoplasmically loading species.
Figures



Similar articles
-
Sieve element occlusion: Interactions with phloem sap-feeding insects. A review.J Plant Physiol. 2022 Feb;269:153582. doi: 10.1016/j.jplph.2021.153582. Epub 2021 Dec 5. J Plant Physiol. 2022. PMID: 34953413 Review.
-
The plant axis as the command centre for (re)distribution of sucrose and amino acids.J Plant Physiol. 2021 Oct;265:153488. doi: 10.1016/j.jplph.2021.153488. Epub 2021 Aug 4. J Plant Physiol. 2021. PMID: 34416599 Review.
-
AtSWEET4, a hexose facilitator, mediates sugar transport to axial sinks and affects plant development.Sci Rep. 2016 Apr 22;6:24563. doi: 10.1038/srep24563. Sci Rep. 2016. PMID: 27102826 Free PMC article.
-
Electrophysiological approach to determine kinetic parameters of sucrose uptake by single sieve elements or phloem parenchyma cells in intact Vicia faba plants.Front Plant Sci. 2013 Jul 31;4:274. doi: 10.3389/fpls.2013.00274. eCollection 2013. Front Plant Sci. 2013. PMID: 23914194 Free PMC article.
-
Phloem loading--not metaphysical, only complex: towards a unified model of phloem loading.J Exp Bot. 1996 Aug;47 Spec No:1155-64. doi: 10.1093/jxb/47.Special_Issue.1155. J Exp Bot. 1996. PMID: 21245244
Cited by
-
Reactive Oxygen Species Initiate Defence Responses of Potato Photosystem II to Sap-Sucking Insect Feeding.Insects. 2022 Apr 24;13(5):409. doi: 10.3390/insects13050409. Insects. 2022. PMID: 35621745 Free PMC article.
-
Phloem cell responses to the feeding activity of Eriosoma lanigerum on Malus domestica.Front Plant Sci. 2025 Jan 30;16:1507552. doi: 10.3389/fpls.2025.1507552. eCollection 2025. Front Plant Sci. 2025. PMID: 39949409 Free PMC article.
-
Involvement of the sieve element cytoskeleton in electrical responses to cold shocks.Plant Physiol. 2013 Jun;162(2):707-19. doi: 10.1104/pp.113.216218. Epub 2013 Apr 26. Plant Physiol. 2013. PMID: 23624858 Free PMC article.
-
Analysis of companion cell and phloem metabolism using a transcriptome-guided model of Arabidopsis metabolism.Plant Physiol. 2023 May 31;192(2):1359-1377. doi: 10.1093/plphys/kiad154. Plant Physiol. 2023. PMID: 36913519 Free PMC article.
-
Characterization of AgMaT2, a plasma membrane mannitol transporter from celery, expressed in phloem cells, including phloem parenchyma cells.Plant Physiol. 2007 Sep;145(1):62-74. doi: 10.1104/pp.107.103143. Epub 2007 Jul 13. Plant Physiol. 2007. PMID: 17631523 Free PMC article.
References
-
- Bertl A, Blumwald E, Coronado R, Eisenberg R, Findlay G, Gradmann D, Hille B, Kohler K, Kolb HA, MacRobbie E, et al (1992) Electrical measurements on endomembranes. Science 258: 873–874 - PubMed
-
- Bethman B, Simonis W, Schönknecht G (1998) Light induced changes of cytosolic pH in Eremosphaera viridis: recordings and kinetic analysis. J Exp Bot 49: 1129–1137
-
- Doering-Saad C, Newbury HJ, Bale JS, Prichard J (2002) Use of aphid stylectomy and RT-PCR for the detection of transporter mRNAs in sieve elements. J Exp Bot 53: 631–637 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
