

The L74V mutation in human immunodeficiency virus type 1 reverse transcriptase counteracts enhanced excision of zidovudine monophosphate associated with thymidine analog resistance mutations
- PMID: 15980332
- PMCID: PMC1168711
- DOI: 10.1128/AAC.49.7.2648-2656.2005
The L74V mutation in human immunodeficiency virus type 1 reverse transcriptase counteracts enhanced excision of zidovudine monophosphate associated with thymidine analog resistance mutations
Abstract
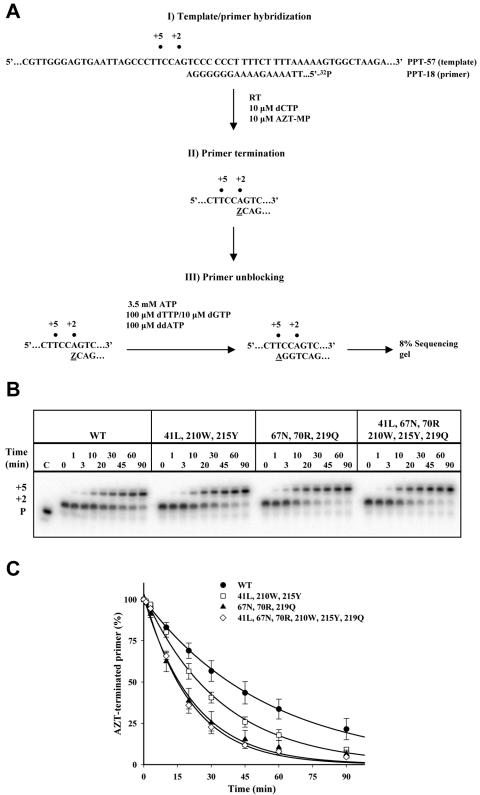
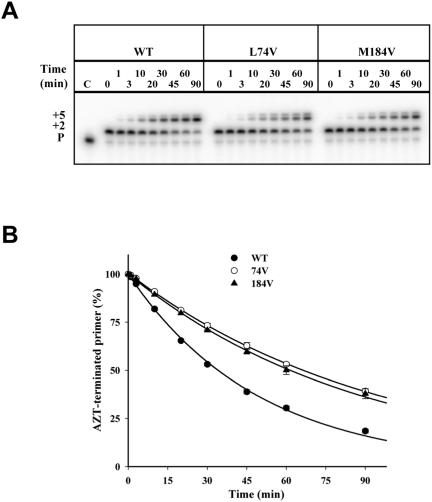
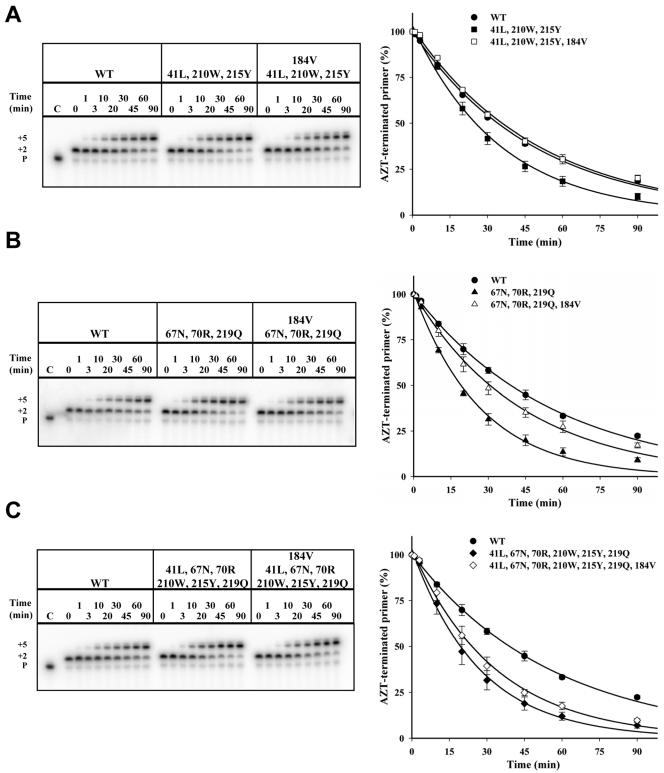
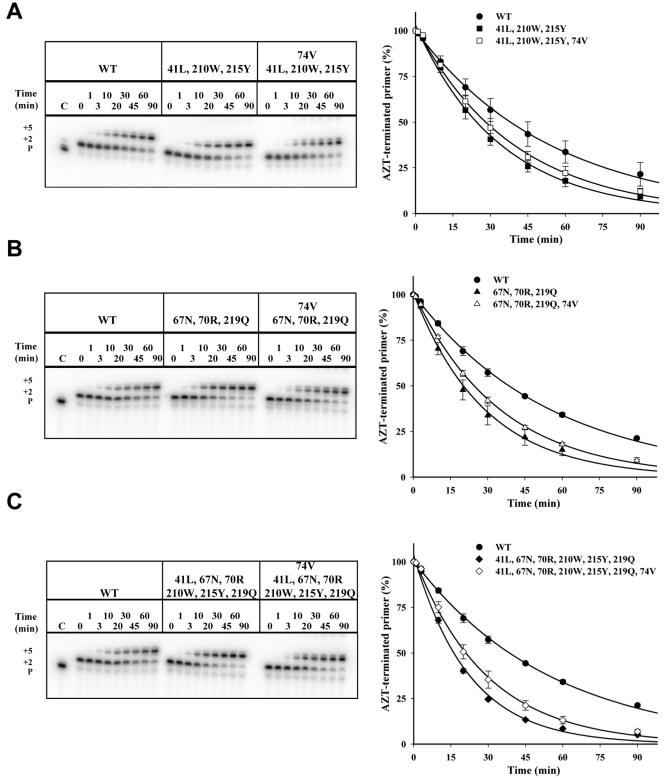
Thymidine analog mutations (TAMs) in human immunodeficiency virus type 1 (HIV-1) reverse transcriptase (RT) confer resistance to zidovudine (AZT) by increasing the rate of ATP-dependent phosphorolysis of the terminal nucleotide monophosphate (primer unblocking). By contrast, the L74V mutation, which confers resistance to didanosine, sensitizes HIV-1 to AZT and partially restores AZT susceptibility when present together with one or more TAMs. To compare rates of primer unblocking in RTs carrying different clusters of TAMs and to explore the biochemical mechanism by which L74V affects AZT susceptibility, ATP-mediated rescue of AZT-blocked DNA synthesis was assayed using a series of purified recombinant RTs. Rates of primer unblocking were higher in the 67N/70R/219Q RT than in the 41L/210W/215Y enzyme and were similar to rates observed with an RT carrying six TAMs (41L/67N/70R/210W/215Y/219Q). The presence of 74V in an otherwise wild-type RT reduced the rate of primer unblocking to a degree similar to that observed with the M184V mutation for lamivudine resistance, which also sensitizes HIV-1 to AZT. Introduction of 74V into RTs carrying TAMs partially counteracted the effect of TAMs on the rate of primer unblocking. The effect of 74V was less marked than that of the 184V mutation in the 67N/70R/219Q and 41L/210W/215Y RTs but similar in the RT carrying six TAMs. These results demonstrate that L74V enhances AZT susceptibility by reducing the extent of its removal by ATP-dependent phosphorolysis and provides further evidence for a common mechanism by which mutations conferring resistance to didanosine and lamivudine sensitize HIV-1 to AZT.
Figures
Similar articles
-
Predictive factors and selection of thymidine analogue mutations by nucleoside reverse transcriptase inhibitors according to initial regimen received.Antivir Ther. 2003 Feb;8(1):65-72. Antivir Ther. 2003. PMID: 12713066 Clinical Trial.
-
The contribution of HIV fitness to the evolution pattern of reverse transcriptase inhibitor resistance.J Med Virol. 2006 Apr;78(4):425-30. doi: 10.1002/jmv.20557. J Med Virol. 2006. PMID: 16482556
-
Impaired rescue of chain-terminated DNA synthesis associated with the L74V mutation in human immunodeficiency virus type 1 reverse transcriptase.Antimicrob Agents Chemother. 2005 Jul;49(7):2657-64. doi: 10.1128/AAC.49.7.2657-2664.2005. Antimicrob Agents Chemother. 2005. PMID: 15980333 Free PMC article.
-
Primer unblocking by HIV-1 reverse transcriptase and resistance to nucleoside RT inhibitors (NRTIs).Int J Biochem Cell Biol. 2004 Sep;36(9):1687-705. doi: 10.1016/j.biocel.2004.02.028. Int J Biochem Cell Biol. 2004. PMID: 15183338 Review.
-
Mutational patterns in the HIV genome and cross-resistance following nucleoside and nucleotide analogue drug exposure.Antivir Ther. 2001;6 Suppl 3:25-44. Antivir Ther. 2001. PMID: 11678471 Review.
Cited by
-
Baseline resistance to nucleoside reverse transcriptase inhibitors fails to predict virologic response to combination therapy in children (PACTG 338).AIDS Res Ther. 2007 Feb 6;4:2. doi: 10.1186/1742-6405-4-2. AIDS Res Ther. 2007. PMID: 17280617 Free PMC article.
-
Variations in reverse transcriptase and RNase H domain mutations in human immunodeficiency virus type 1 clinical isolates are associated with divergent phenotypic resistance to zidovudine.Antimicrob Agents Chemother. 2007 Nov;51(11):3861-9. doi: 10.1128/AAC.00646-07. Epub 2007 Aug 27. Antimicrob Agents Chemother. 2007. PMID: 17724152 Free PMC article.
-
The "Connection" Between HIV Drug Resistance and RNase H.Viruses. 2010 Jul 1;2(7):1476-1503. doi: 10.3390/v2071476. Viruses. 2010. PMID: 21088701 Free PMC article.
-
HIV-1 reverse transcriptase (RT) polymorphism 172K suppresses the effect of clinically relevant drug resistance mutations to both nucleoside and non-nucleoside RT inhibitors.J Biol Chem. 2012 Aug 24;287(35):29988-99. doi: 10.1074/jbc.M112.351551. Epub 2012 Jul 2. J Biol Chem. 2012. PMID: 22761416 Free PMC article.
-
Mutations in the connection domain of HIV-1 reverse transcriptase increase 3'-azido-3'-deoxythymidine resistance.Proc Natl Acad Sci U S A. 2007 Jan 2;104(1):317-22. doi: 10.1073/pnas.0609642104. Epub 2006 Dec 19. Proc Natl Acad Sci U S A. 2007. PMID: 17179211 Free PMC article.
References
-
- Arion, D., N. Kaushik, S. McCormick, G. Borkow, and M. Parniak. 1998. Phenotypic mechanism of HIV-1 resistance to 3′-azido-3′-deoxythymidine (AZT): increased polymerization processivity and enhanced sensitivity to pyrophosphate of the mutant viral reverse transcriptase. Biochemistry 37:15908-15917. - PubMed
-
- Back, N. K. T., M. Nijhuis, W. Keulen, C. A. B. Boucher, B. O. Oude Essink, A. B. van Kuilenburg, A. H. van Gennip, and B. Berkhout. 1996. Reduced replication of 3TC-resistant HIV-1 variants in primary cells due to a processivity defect of the reverse transcriptase enzyme. EMBO J. 15:4040-4049. - PMC - PubMed
-
- Boucher, C. A. B., E. O'Sullivan, J. W. Mulder, C. Ramautarsing, P. Kellam, G. Darby, J. M. A. Lange, J. Goudsmit, and B. A. Larder. 1992. Ordered appearance of zidovudine resistance mutations during treatment of 18 human immunodeficiency virus-positive subjects. J. Infect. Dis. 165:105-110. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
