

The hypoglycaemic activity of fenugreek seed extract is mediated through the stimulation of an insulin signalling pathway
- PMID: 15980869
- PMCID: PMC1576255
- DOI: 10.1038/sj.bjp.0706312
The hypoglycaemic activity of fenugreek seed extract is mediated through the stimulation of an insulin signalling pathway
Abstract
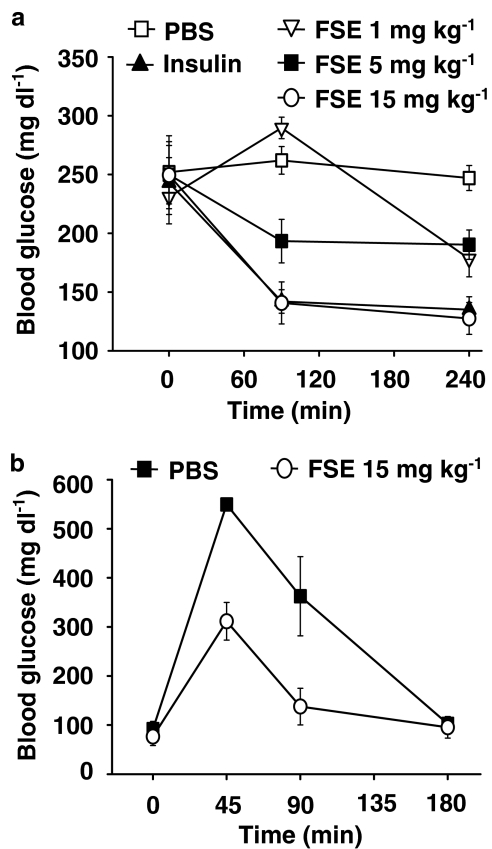
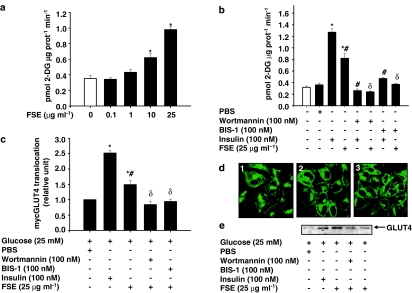
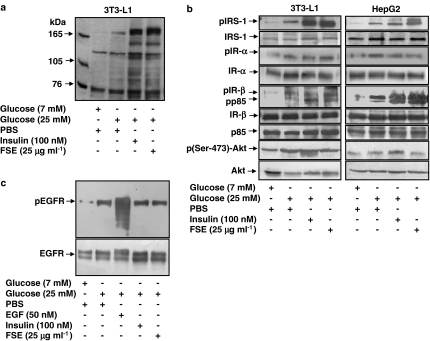
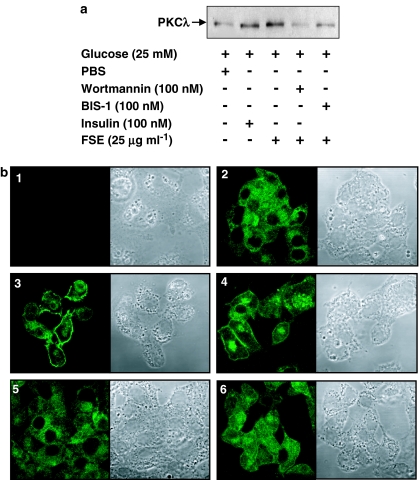
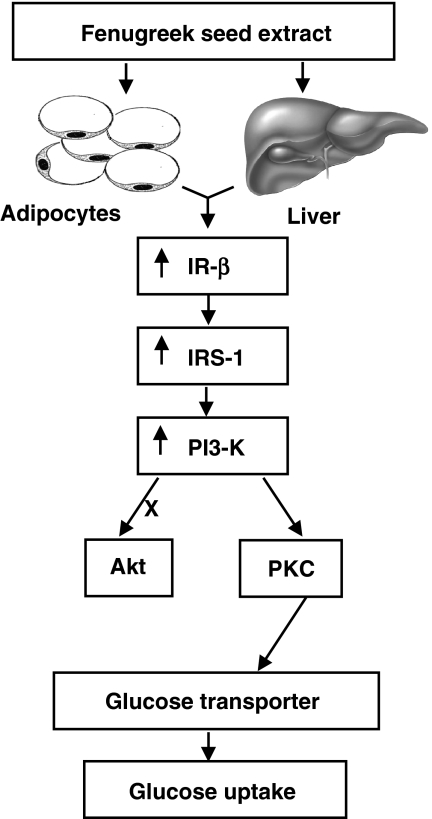
The in vivo hypoglycaemic activity of a dialysed fenugreek seed extract (FSE) was studied in alloxan (AXN)-induced diabetic mice and found to be comparable to that of insulin (1.5 U kg(-1)). FSE also improved intraperitoneal glucose tolerance in normal mice. The mechanism by which FSE attenuated hyperglycaemia was investigated in vitro. FSE stimulated glucose uptake in CHO-HIRc-mycGLUT4eGFP cells in a dose-dependent manner. This effect was shown to be mediated by the translocation of glucose transporter 4 (GLUT4) from the intracellular space to the plasma membrane. These effects of FSE on GLUT4 translocation and glucose uptake were inhibited by wortmannin, a phosphatidylinositol 3-kinase (PI3-K) inhibitor, and bisindolylmaleimide 1, a protein kinase C (PKC)-specific inhibitor. In vitro phosphorylation analysis revealed that, like insulin, FSE also induces tyrosine phosphorylation of a number of proteins including the insulin receptor, insulin receptor substrate 1 and p85 subunit of PI3-K, in both 3T3-L1 adipocytes and human hepatoma cells, HepG2. However, unlike insulin, FSE had no effect on protein kinase B (Akt) activation. These results suggest that in vivo the hypoglycaemic effect of FSE is mediated, at least in part, by the activation of an insulin signalling pathway in adipocytes and liver cells.
Figures
References
-
- ABDEL-BARRY J.A., ABDEL-HASSAN I.A., AL-HAKIEM M.H. Hypoglycemic and antihyperglycemic effects of Trigonella foenum-graecum leaf in normal and alloxan induced diabetic rats. J. Ethnopharmacol. 1997;58:149–155. - PubMed
-
- BASCH E., ULBRICHT C., KUO G., SZAPARY P., SMITH M. Therapeutic applications of fenugreek. Altern. Med. Rev. 2003;8:20–27. - PubMed
-
- CHHIPA R.R., SINGH S., SURVE S.V., VIJAYAKUMAR M.V., BHAT M.K. Doxycycline potentiates antitumor effect of cyclophosphamide in mice. Toxicol. Appl. Pharmacol. 2005;202:268–277. - PubMed
-
- CLANCY B.M., CZECH M.P. Hexose transport stimulation and membrane redistribution of glucose transporter isoforms in response to cholera toxin, dibutyryl cyclic AMP, and insulin in 3T3-L1 adipocytes. J. Biol. Chem. 1990;265:12434–12443. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
