

A potential link between transgene silencing and poly(A) tails
- PMID: 15987811
- PMCID: PMC1370785
- DOI: 10.1261/rna.2280105
A potential link between transgene silencing and poly(A) tails
Abstract
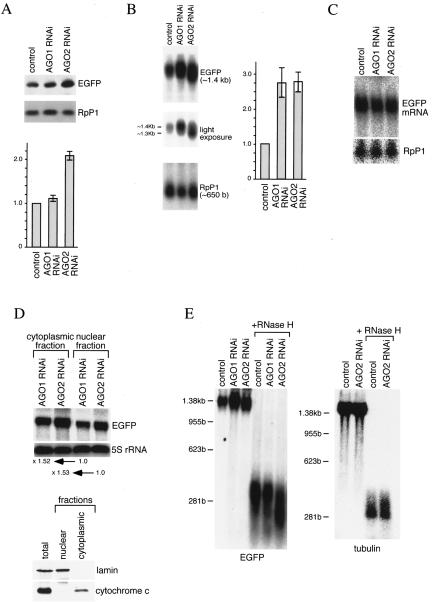
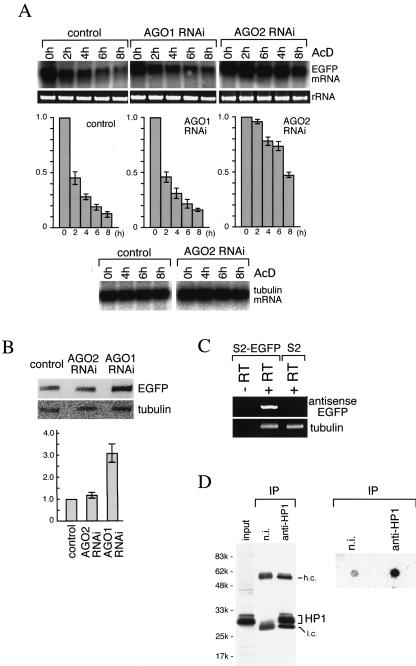
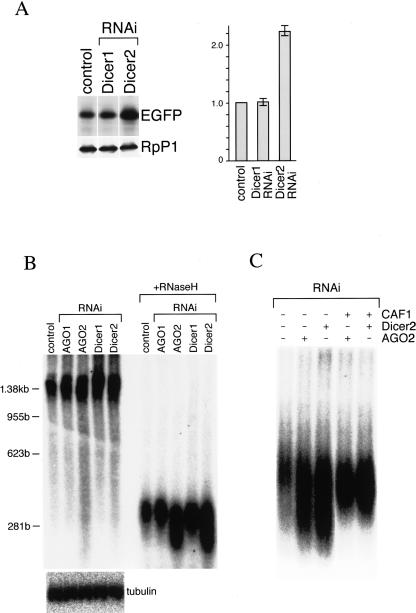
Argonaute proteins function in gene silencing induced by double-stranded RNA (dsRNA) in various organisms. In Drosophila, the Argonaute proteins AGO1 and AGO2 have been implicated in post-transcriptional gene-silencing (PTGS)/RNA interference (RNAi). In this study, we found that AGO1 and AGO2 depletion caused the accumulation of multicopied enhanced green fluorescence protein (EGFP) transgene transcripts in Drosophila S2 cells. Depletion of AGO1, the essential factor for miRNA biogenesis, led to an increased transcriptional rate of the transgenes. In contrast, depletion of AGO2, the essential factor for siRNA-directed RNAi, resulted in EGFP mRNA stabilization with concomitant shortening of the EGFP mRNA poly(A) tail. Our findings suggest that AGO1 and AGO2 mediate multicopied transgene silencing by different mechanisms. Intriguingly, Dicer2 depletion phenocopies AGO2 depletion, with an increase in EGFP protein levels and shortening of the EGFP mRNA poly(A) tail. The possibility that AGO2 and Dicer2 involve, at least in part, poly(A) length maintenance of transgene mRNA suggests a potentially important link between transgene silencing and poly(A) tails.
Figures
Similar articles
-
Distinct roles for Argonaute proteins in small RNA-directed RNA cleavage pathways.Genes Dev. 2004 Jul 15;18(14):1655-66. doi: 10.1101/gad.1210204. Epub 2004 Jul 1. Genes Dev. 2004. PMID: 15231716 Free PMC article.
-
Genome-wide analysis of mRNAs regulated by Drosha and Argonaute proteins in Drosophila melanogaster.Mol Cell Biol. 2006 Apr;26(8):2965-75. doi: 10.1128/MCB.26.8.2965-2975.2006. Mol Cell Biol. 2006. PMID: 16581772 Free PMC article.
-
Argonaute-mediated translational repression (and activation).Fly (Austin). 2009 Jul-Sep;3(3):204-6. Epub 2009 Jul 14. Fly (Austin). 2009. PMID: 19556851 Review.
-
Sorting of Drosophila small silencing RNAs partitions microRNA* strands into the RNA interference pathway.RNA. 2010 Jan;16(1):43-56. doi: 10.1261/rna.1972910. Epub 2009 Nov 16. RNA. 2010. PMID: 19917635 Free PMC article.
-
MicroRNAs silence gene expression by repressing protein expression and/or by promoting mRNA decay.Cold Spring Harb Symp Quant Biol. 2006;71:523-30. doi: 10.1101/sqb.2006.71.013. Cold Spring Harb Symp Quant Biol. 2006. PMID: 17381335 Review.
Cited by
-
New construct approaches for efficient gene silencing in plants.Plant Physiol. 2006 Aug;141(4):1508-18. doi: 10.1104/pp.106.082271. Epub 2006 Jun 9. Plant Physiol. 2006. PMID: 16766670 Free PMC article.
-
Sensitive detection of GFP utilizing tyramide signal amplification to overcome gene silencing.Exp Cell Res. 2007 May 15;313(9):1943-50. doi: 10.1016/j.yexcr.2007.02.024. Epub 2007 Mar 12. Exp Cell Res. 2007. PMID: 17428464 Free PMC article.
-
RNA helicase Belle/DDX3 regulates transgene expression in Drosophila.Dev Biol. 2016 Apr 1;412(1):57-70. doi: 10.1016/j.ydbio.2016.02.014. Epub 2016 Feb 18. Dev Biol. 2016. PMID: 26900887 Free PMC article.
-
Slicer function of Drosophila Argonautes and its involvement in RISC formation.Genes Dev. 2005 Dec 1;19(23):2837-48. doi: 10.1101/gad.1370605. Epub 2005 Nov 14. Genes Dev. 2005. PMID: 16287716 Free PMC article.
-
Basic concepts of epigenetics: impact of environmental signals on gene expression.Epigenetics. 2012 Feb;7(2):119-30. doi: 10.4161/epi.7.2.18764. Epigenetics. 2012. PMID: 22395460 Free PMC article. Review.
References
-
- Ambros, V. 2004. The function of animal microRNAs. Nature 431: 350–355. - PubMed
-
- Aravin, A.A., Naumova, N.M., Tulin, A.V., Vagin, V.V., Rozovsky, Y.M., and Gvozdev, V.A. 2001. Double-stranded RNA-mediated silencing of genomic tandem repeats and transposable elements in the D. melanogaster germline. Curr. Biol. 11: 1017–1027. - PubMed
-
- Bartel, D.P. 2004. MicroRNAs: Genomics, biogenesis, mechanism, and function. Cell 116: 281–297. - PubMed
-
- Baulcombe, D. 2004. RNA silencing in plants. Nature 431: 356–363. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases