

Reciprocal transcriptional regulation of Pou5f1 and Sox2 via the Oct4/Sox2 complex in embryonic stem cells
- PMID: 15988017
- PMCID: PMC1168830
- DOI: 10.1128/MCB.25.14.6031-6046.2005
Reciprocal transcriptional regulation of Pou5f1 and Sox2 via the Oct4/Sox2 complex in embryonic stem cells
Abstract
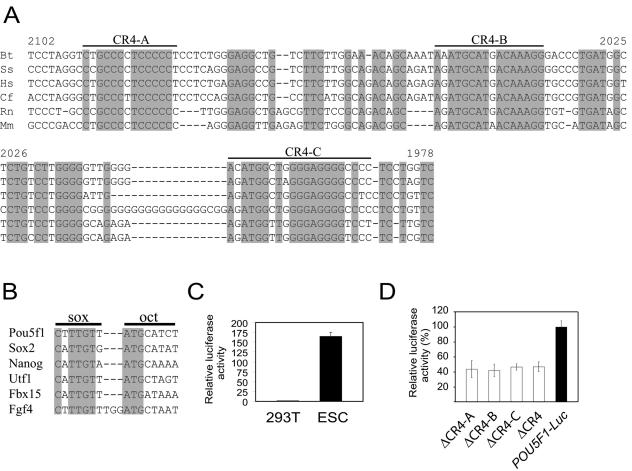
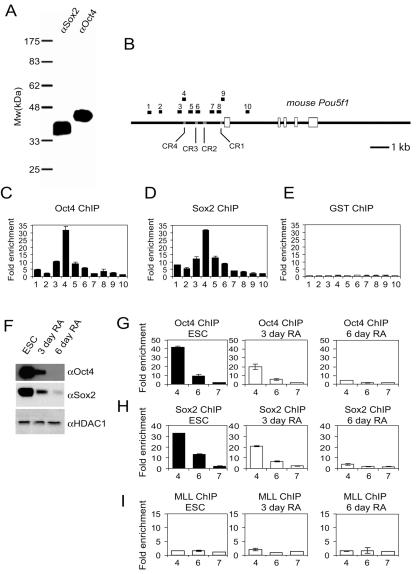
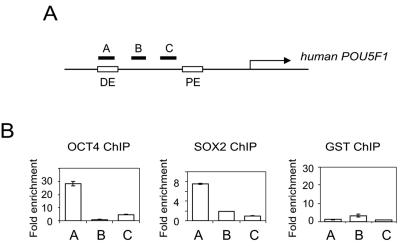
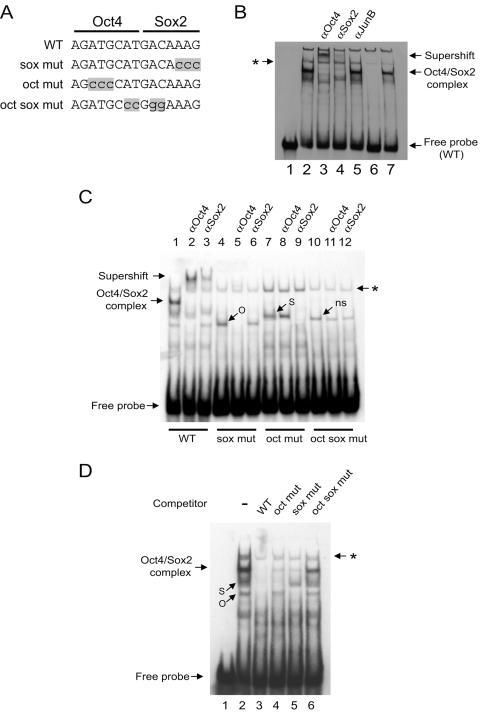
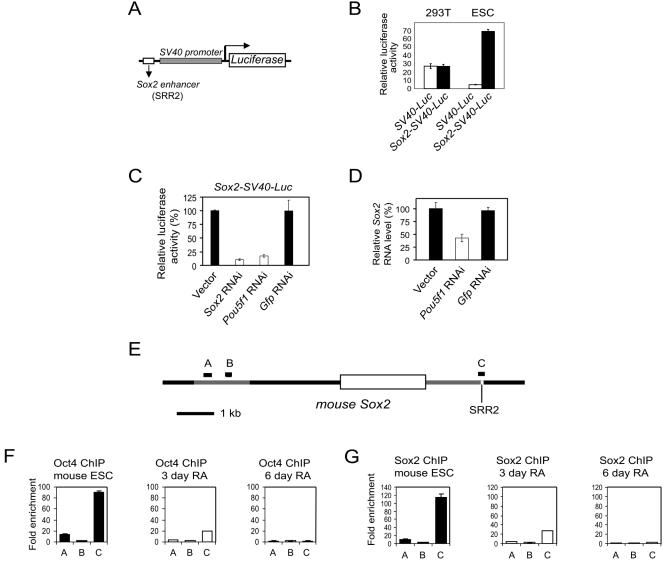
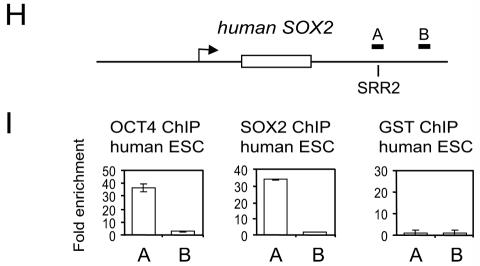
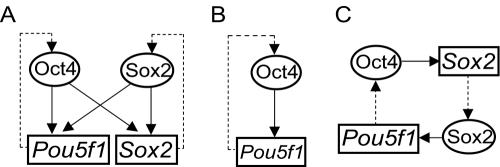
Embryonic stem cells (ESCs) are pluripotent cells that can either self-renew or differentiate into many cell types. Oct4 and Sox2 are transcription factors essential to the pluripotent and self-renewing phenotypes of ESCs. Both factors are upstream in the hierarchy of the transcription regulatory network and are partners in regulating several ESC-specific genes. In ESCs, Sox2 is transcriptionally regulated by an enhancer containing a composite sox-oct element that Oct4 and Sox2 bind in a combinatorial interaction. It has previously been shown that Pou5f1, the Oct4 gene, contains a distal enhancer imparting specific expression in both ESCs and preimplantation embryos. Here, we identify a composite sox-oct element within this enhancer and show that it is involved in Pou5f1 transcriptional activity in ESCs. In vitro experiments with ESC nuclear extracts demonstrate that Oct4 and Sox2 interact specifically with this regulatory element. More importantly, by chromatin immunoprecipitation assay, we establish that both Oct4 and Sox2 bind directly to the composite sox-oct elements in both Pou5f1 and Sox2 in living mouse and human ESCs. Specific knockdown of either Oct4 or Sox2 by RNA interference leads to the reduction of both genes' enhancer activities and endogenous expression levels in addition to ESC differentiation. Our data uncover a positive and potentially self-reinforcing regulatory loop that maintains Pou5f1 and Sox2 expression via the Oct4/Sox2 complex in pluripotent cells.
Figures
References
-
- Ambrosetti, D. C., H. R. Scholer, L. Dailey, and C. Basilico. 2000. Modulation of the activity of multiple transcriptional activation domains by the DNA binding domains mediates the synergistic action of Sox2 and Oct-3 on the fibroblast growth factor-4 enhancer. J. Biol. Chem. 275:23387-23397. - PubMed
-
- Ben-Shushan, E., J. R. Thompson, L. J. Gudas, and Y. Bergman. 1998. Rex-1, a gene encoding a transcription factor expressed in the early embryo, is regulated via Oct-3/4 and Oct-6 binding to an octamer site and a novel protein, Rox-1, binding to an adjacent site. Mol. Cell. Biol. 18:1866-1878. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous