

Acyl-coenzyme A:cholesterol acyltransferase promotes oxidized LDL/oxysterol-induced apoptosis in macrophages
- PMID: 15995174
- PMCID: PMC2768430
- DOI: 10.1194/jlr.M500101-JLR200
Acyl-coenzyme A:cholesterol acyltransferase promotes oxidized LDL/oxysterol-induced apoptosis in macrophages
Abstract
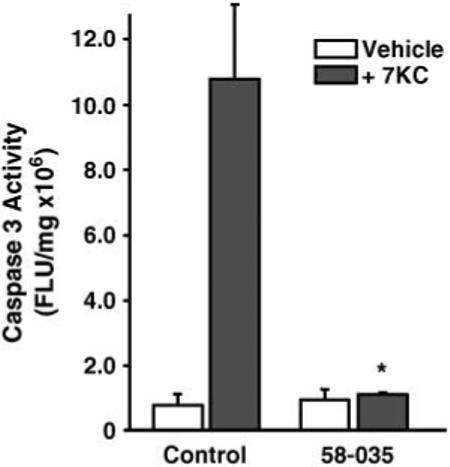
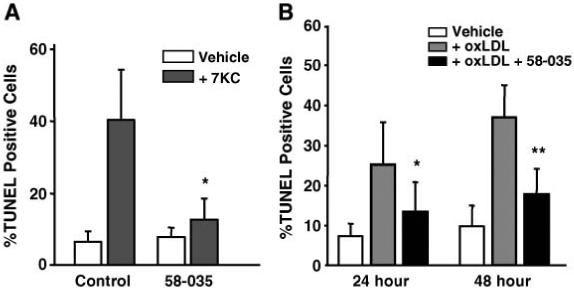
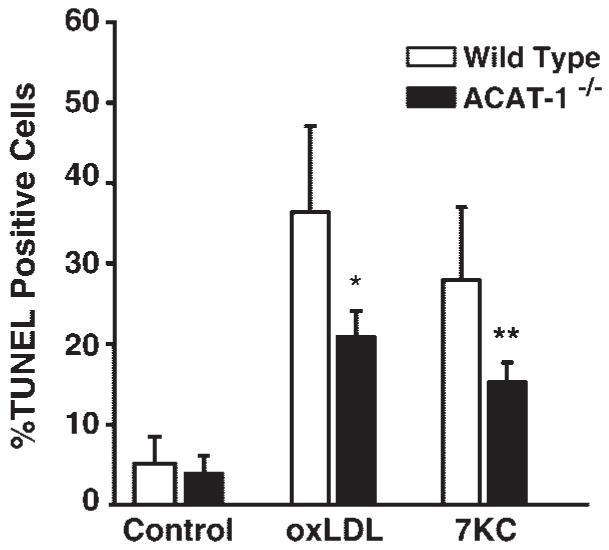
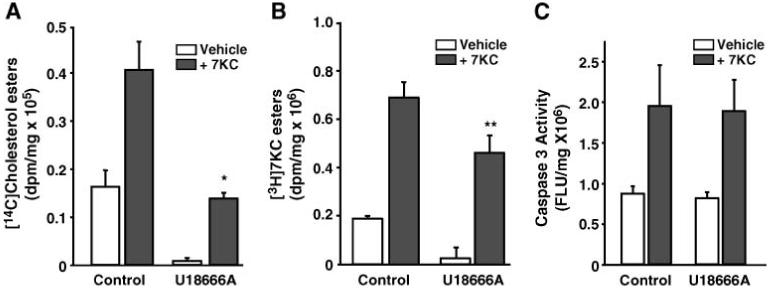
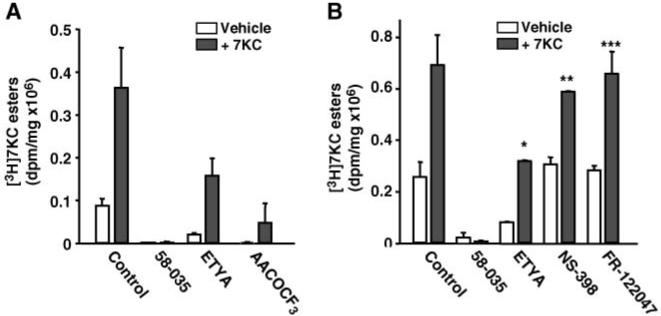
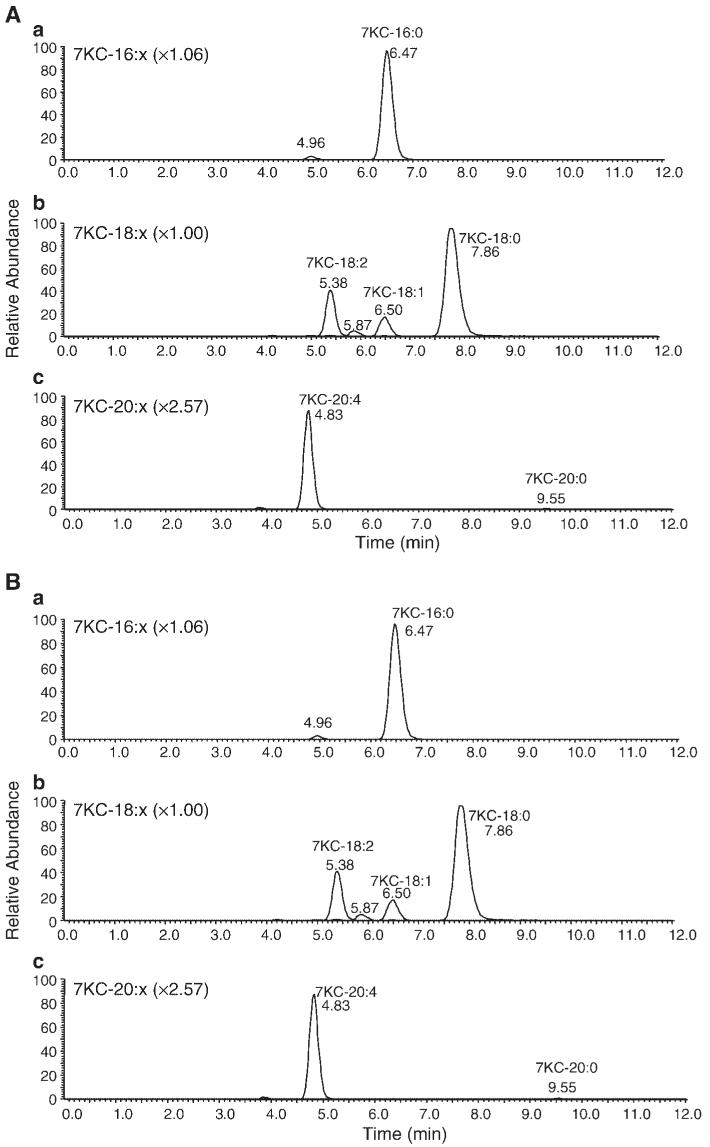
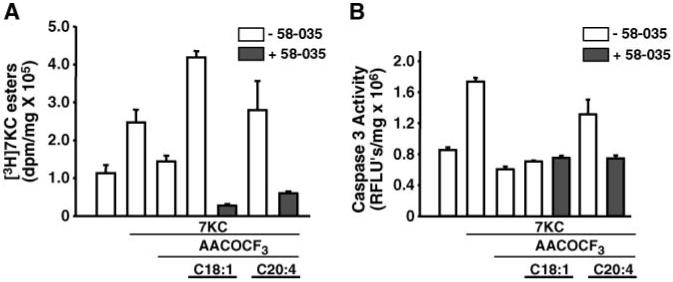
7-Ketocholesterol (7KC) is a cytotoxic component of oxidized low density lipoproteins (OxLDLs) and induces apoptosis in macrophages by a mechanism involving the activation of cytosolic phospholipase A2 (cPLA2). In the current study, we examined the role of ACAT in 7KC-induced and OxLDL-induced apoptosis in murine macrophages. An ACAT inhibitor, Sandoz 58-035, suppressed 7KC-induced apoptosis in P388D1 cells and both 7KC-induced and OxLDL-induced apoptosis in mouse peritoneal macrophages (MPMs). Furthermore, compared with wild-type MPMs, ACAT-1-deficient MPMs demonstrated significant resistance to both 7KC-induced and OxLDL-induced apoptosis. Macrophages treated with 7KC accumulated ACAT-derived [14C]cholesteryl and [3H]7-ketocholesteryl esters. Tandem LC-MS revealed that the 7KC esters contained primarily saturated and monounsaturated fatty acids. An inhibitor of cPLA2, arachidonyl trifluoromethyl ketone, prevented the accumulation of 7KC esters and inhibited 7KC-induced apoptosis in P388D1 cells. The decrease in 7KC ester accumulation produced by the inhibition of cPLA2 was reversed by supplementing with either oleic or arachidonic acid (AA); however, only AA supplementation restored the induction of apoptosis by 7KC. These results suggest that 7KC not only initiates the apoptosis pathway by activating cPLA2, as we have reported previously, but also participates in the downstream signaling pathway when esterified by ACAT to form 7KC-arachidonate.
Figures
Similar articles
-
Cholesterol and oxysterol metabolism and subcellular distribution in macrophage foam cells. Accumulation of oxidized esters in lysosomes.J Lipid Res. 2000 Feb;41(2):226-37. J Lipid Res. 2000. PMID: 10681406
-
Oxidized LDL activates phospholipase A2 to supply fatty acids required for cholesterol esterification.J Lipid Res. 2003 Sep;44(9):1676-85. doi: 10.1194/jlr.M300012-JLR200. Epub 2003 Jun 1. J Lipid Res. 2003. PMID: 12777475
-
[Involvement of phospholipase A2 in the supply of fatty acids required for cholesterol esterification associated with uptake of oxidized low-density lipoprotein in macrophages].Yakugaku Zasshi. 2003 Oct;123(10):845-53. doi: 10.1248/yakushi.123.845. Yakugaku Zasshi. 2003. PMID: 14577330 Review. Japanese.
-
Arachidonate metabolism and the signaling pathway of induction of apoptosis by oxidized LDL/oxysterol.J Lipid Res. 2001 Oct;42(10):1678-86. J Lipid Res. 2001. PMID: 11590225
-
Human acyl-CoA:cholesterol acyltransferase (ACAT) and its potential as a target for pharmaceutical intervention against atherosclerosis.Acta Biochim Biophys Sin (Shanghai). 2006 Mar;38(3):151-6. doi: 10.1111/j.1745-7270.2006.00154.x. Acta Biochim Biophys Sin (Shanghai). 2006. PMID: 16518538 Review.
Cited by
-
Cannabinoid (CB2) receptor deficiency reduces the susceptibility of macrophages to oxidized LDL/oxysterol-induced apoptosis.J Lipid Res. 2008 Nov;49(11):2338-46. doi: 10.1194/jlr.M800105-JLR200. Epub 2008 Jul 9. J Lipid Res. 2008. PMID: 18614816 Free PMC article.
-
11β-Hydroxysteroid dehydrogenase type 1 contributes to the regulation of 7-oxysterol levels in the arterial wall through the inter-conversion of 7-ketocholesterol and 7β-hydroxycholesterol.Biochimie. 2013 Mar;95(3):548-55. doi: 10.1016/j.biochi.2012.08.007. Epub 2012 Aug 23. Biochimie. 2013. PMID: 22940536 Free PMC article.
-
Rimonabant is a dual inhibitor of acyl CoA:cholesterol acyltransferases 1 and 2.Biochem Biophys Res Commun. 2010 Aug 6;398(4):671-6. doi: 10.1016/j.bbrc.2010.06.134. Epub 2010 Jul 6. Biochem Biophys Res Commun. 2010. PMID: 20609360 Free PMC article.
-
11β-Hydroxysteroid dehydrogenase type 1 contributes to the balance between 7-keto- and 7-hydroxy-oxysterols in vivo.Biochem Pharmacol. 2013 Jul 1;86(1):146-53. doi: 10.1016/j.bcp.2013.02.002. Epub 2013 Feb 13. Biochem Pharmacol. 2013. PMID: 23415904 Free PMC article.
-
7-Ketocholesterol Induces Lipid Metabolic Reprogramming and Enhances Cholesterol Ester Accumulation in Cardiac Cells.Cells. 2021 Dec 20;10(12):3597. doi: 10.3390/cells10123597. Cells. 2021. PMID: 34944104 Free PMC article.
References
-
- Ball RY, Stowers EC, Burton JH, Cary NR, Skepper JN, Mitchinson MJ. Evidence that the death of macrophage foam cells contributes to the lipid core of atheroma. Atherosclerosis. 1995;114:45–54. - PubMed
-
- Colles SM, Maxson JM, Carlson SG, Chisolm GM. Oxidized LDL-induced injury and apoptosis in arthrosclerosis. Potential roles for oxysterols. Trends Cardiovasc. Med. 2001;11:131–138. - PubMed
-
- Yao PM, Tabas I. Free cholesterol loading of macrophages induces apoptosis involving the Fas pathway. J. Biol. Chem. 2000;275:23807–23818. - PubMed
-
- Steinberg D, Witztum JL. Is the oxidative modification hypothesis relevant to human atherosclerosis? Circulation. 2002;105:2107–2111. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
