

The Burkholderia pseudomallei BpeAB-OprB efflux pump: expression and impact on quorum sensing and virulence
- PMID: 15995185
- PMCID: PMC1169529
- DOI: 10.1128/JB.187.14.4707-4719.2005
The Burkholderia pseudomallei BpeAB-OprB efflux pump: expression and impact on quorum sensing and virulence
Abstract
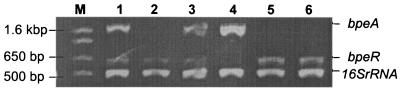
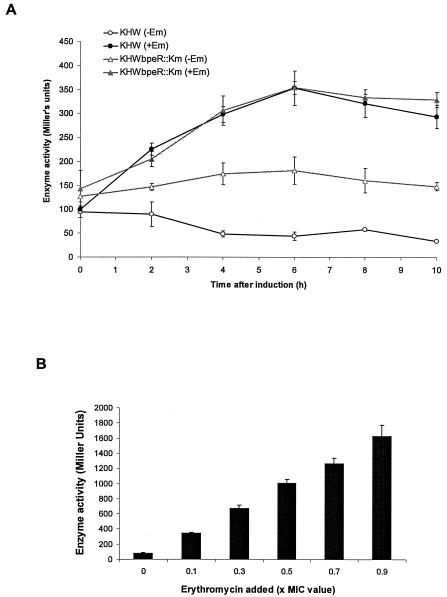
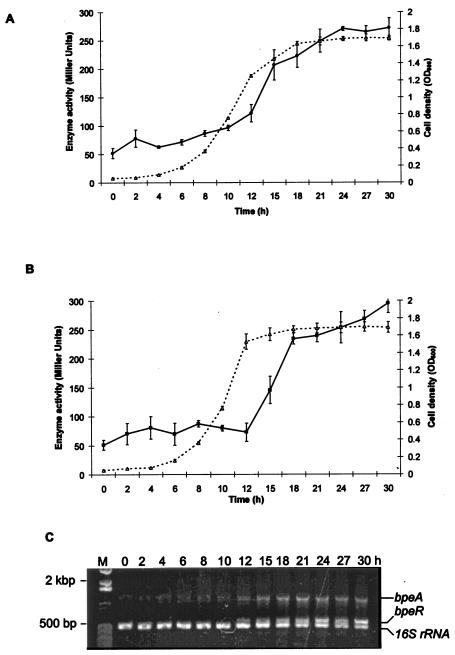
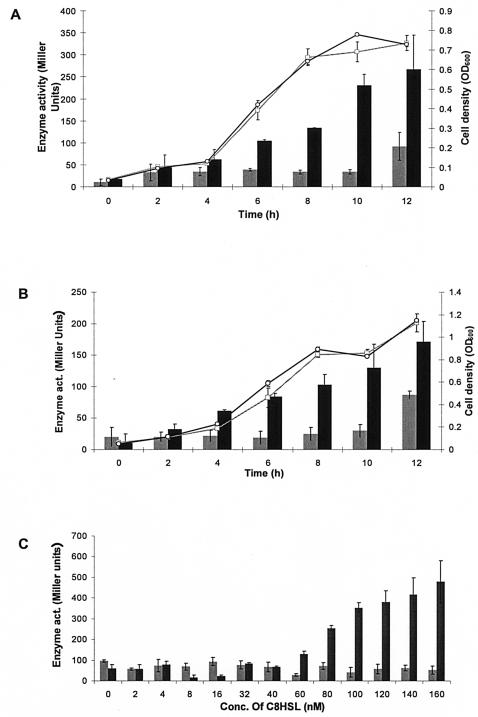
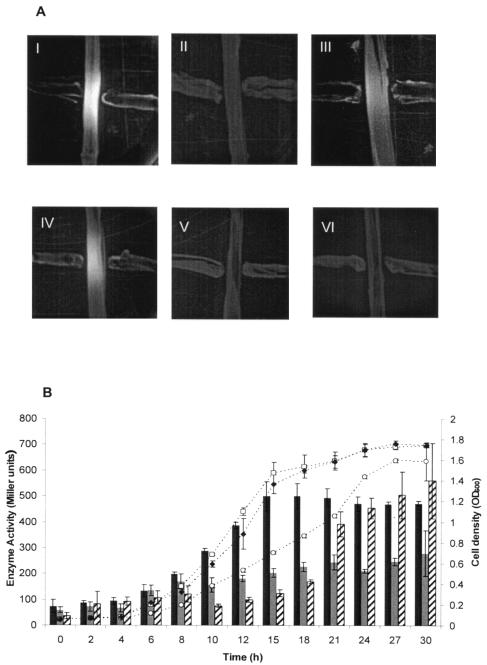
BpeAB-OprB is a multidrug efflux pump of the bacterial pathogen Burkholderia pseudomallei and is responsible for conferring antimicrobial resistance to aminoglycosides and macrolides. Expression of bpeAB-oprB is inducible by its substrate erythromycin and upon entry into stationary phase. BpeR, a member of the TetR family, functions as a repressor of the bpeAB-oprB operon. bpeR expression was similarly induced at stationary phase but lagged behind the induction of bpeAB-oprB expression. The induction of bpeAB-oprB expression could be advanced to the early exponential phase by exogenous addition of the B. pseudomallei autoinducers N-octanoyl-homoserine lactone (C8HSL) and N-decanoyl-homoserine lactone (C10HSL), suggesting that the bpeAB-oprB operon may be quorum regulated. On the other hand, acyl-homoserine lactone (acyl-HSL) production was undetectable in the bpeAB-null mutant and strains which overexpress bpeR. The failure of these strains to produce acyl-HSLs seemed to be at the level of synthesis of acyl-HSLs, as growth-phase-dependent expression of the autoinducer synthase BpsI was abolished in the bpeAB-null mutant. bpsI expression remained growth phase dependent in the bpeR mutant which had functional BpeAB-OprB. BpeAB-OprB function is likewise necessary for optimal production of quorum-sensing-controlled virulence factors such as siderophore and phospholipase C and for biofilm formation. Cell invasion and cytotoxicity towards human lung epithelial (A549) and human macrophage (THP-1) cells were also significantly attenuated in both the bpeAB mutant and bpeR-overexpressing strains, thus suggesting the possibility of attenuating B. pseudomallei virulence using inhibitors of the BpeAB-OprB efflux pump.
Figures
References
-
- Casadaban, M. J., J. Chou, and S. N. Cohen. 1980. In vitro gene fusions that join an enzymatically active beta-galactosidase segment to amino-terminal fragments of exogenous proteins: Escherichia coli plasmid vectors for the detection and cloning of translational initiation signals. J. Bacteriol. 143:971-980. - PMC - PubMed
-
- Chaowagul, W. 2000. Recent advances in the treatment of severe melioidosis. Acta Trop. 74:133-137. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
