

Analysis of the Escherichia coli Alp phenotype: heat shock induction in ssrA mutants
- PMID: 15995188
- PMCID: PMC1169506
- DOI: 10.1128/JB.187.14.4739-4751.2005
Analysis of the Escherichia coli Alp phenotype: heat shock induction in ssrA mutants
Abstract
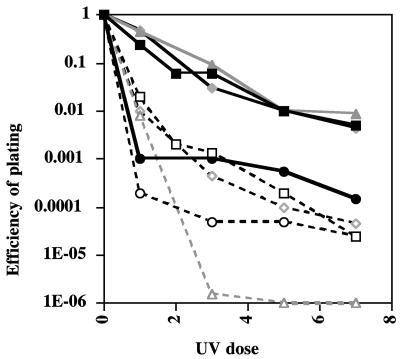
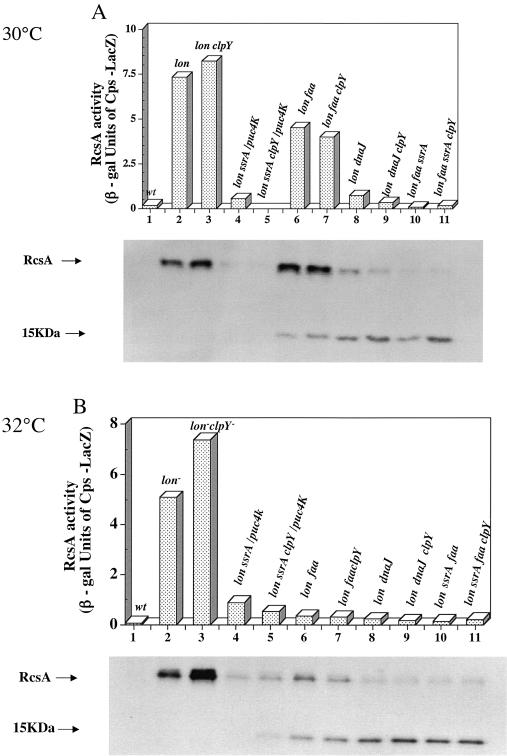
The major phenotypes of lon mutations, UV sensitivity and overproduction of capsule, are due to the stabilization of two substrates, SulA and RcsA. Inactivation of transfer mRNA (tmRNA) (encoded by ssrA), coupled with a multicopy kanamycin resistance determinant, suppressed both lon phenotypes and restored the rapid degradation of SulA. This novel protease activity was named Alp but was never identified further. We report here the identification, mapping, and characterization of a chromosomal mutation, faa (for function affecting Alp), that leads to full suppression of a Deltalon ssrA::cat host and thus bypasses the requirement for multicopy Kan(r); faa and ssrA mutants are additive in their ability to suppress lon mutants. The faa mutation was mapped to the C terminus of dnaJ(G232); dnaJ null mutants have similar effects. The identification of a lon suppressor in dnaJ suggested the possible involvement of heat shock. We find that ssrA mutants alone significantly induce the heat shock response. The suppression of UV sensitivity, both in the original Alp strain and in faa mutants, is reversed by mutations in clpY, encoding a subunit of the heat shock-induced ClpYQ protease that is known to degrade SulA. However, capsule synthesis is not restored by clpY mutants, probably because less RcsA accumulates in the Alp strain and because the RcsA that does accumulate is inactive. Both ssrA effects are partially relieved by ssrA derivatives encoding protease-resistant tags, implicating ribosome stalling as the primary defect. Thus, ssrA and faa each suppress two lon mutant phenotypes but by somewhat different mechanisms, with heat shock induction playing a major role.
Figures





Similar articles
-
Two new mutations in dnaJ suppress DNA damage hypersensitivity and capsule overproduction phenotypes of Δlon mutant of Escherichia coli by modulating the expression of clpYQ (hslUV) and rcsA genes.Gene. 2020 Feb 5;726:144135. doi: 10.1016/j.gene.2019.144135. Epub 2019 Oct 4. Gene. 2020. PMID: 31589958
-
Suppression of Δlon phenotypes in Escherichia coli by N-terminal DnaK peptides.J Basic Microbiol. 2019 Mar;59(3):302-313. doi: 10.1002/jobm.201800469. Epub 2019 Jan 7. J Basic Microbiol. 2019. PMID: 30614541
-
Redundant in vivo proteolytic activities of Escherichia coli Lon and the ClpYQ (HslUV) protease.J Bacteriol. 1999 Jun;181(12):3681-7. doi: 10.1128/JB.181.12.3681-3687.1999. J Bacteriol. 1999. PMID: 10368141 Free PMC article.
-
Alp, a suppressor of lon protease mutants in Escherichia coli.J Bacteriol. 1989 Jun;171(6):3348-53. doi: 10.1128/jb.171.6.3348-3353.1989. J Bacteriol. 1989. PMID: 2656652 Free PMC article.
-
SulA-dependent hypersensitivity to high pressure and hyperfilamentation after high-pressure treatment of Escherichia coli lon mutants.Res Microbiol. 2005 Mar;156(2):233-7. doi: 10.1016/j.resmic.2004.10.003. Epub 2004 Dec 1. Res Microbiol. 2005. PMID: 15748989
Cited by
-
Prophage excision switches the primary ribosome rescue pathway and rescue-associated gene regulations in Escherichia coli.Mol Microbiol. 2023 Jan;119(1):44-58. doi: 10.1111/mmi.15003. Epub 2022 Dec 5. Mol Microbiol. 2023. PMID: 36471624 Free PMC article.
-
A putative curved DNA region upstream of rcsA in Escherichia coli plays a key role in transcriptional regulation by H-NS.FEBS Open Bio. 2018 Jun 26;8(8):1209-1218. doi: 10.1002/2211-5463.12348. eCollection 2018 Aug. FEBS Open Bio. 2018. PMID: 30087827 Free PMC article.
-
Importance of the tmRNA system for cell survival when transcription is blocked by DNA-protein cross-links.Mol Microbiol. 2010 Nov;78(3):686-700. doi: 10.1111/j.1365-2958.2010.07355.x. Epub 2010 Sep 16. Mol Microbiol. 2010. PMID: 20807197 Free PMC article.
-
Characterization of the Escherichia coli ClpY (HslU) substrate recognition site in the ClpYQ (HslUV) protease using the yeast two-hybrid system.J Bacteriol. 2009 Jul;191(13):4218-31. doi: 10.1128/JB.00089-09. Epub 2009 Apr 24. J Bacteriol. 2009. PMID: 19395483 Free PMC article.
-
Small RNAs and small proteins involved in resistance to cell envelope stress and acid shock in Escherichia coli: analysis of a bar-coded mutant collection.J Bacteriol. 2010 Jan;192(1):59-67. doi: 10.1128/JB.00873-09. J Bacteriol. 2010. PMID: 19734312 Free PMC article.
References
-
- Chuang, S.-E., I. G. P. B. Burland, D. L. Daniels, and F. R. Blattner. 1993. Sequence analysis of four new heat-shock genes constituting the hslTS/ibpAB and hslVU operons in Escherichia coli. Gene 134:1-6. - PubMed
-
- Goffin, L., and C. Georgopoulos. 1998. Genetic and biochemical characterization of mutations affecting the carboxy-terminal domain of the Escherichia coli molecular chaperone DnaJ. Mol. Microbiol. 30:329-340. - PubMed
-
- Gottesman, S. 1996. Proteases and their targets in Escherichia coli. Annu. Rev. Genet. 30:465-506. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
