

Amadori adducts activate nuclear factor-kappaB-related proinflammatory genes in cultured human peritoneal mesothelial cells
- PMID: 15997235
- PMCID: PMC1576262
- DOI: 10.1038/sj.bjp.0706309
Amadori adducts activate nuclear factor-kappaB-related proinflammatory genes in cultured human peritoneal mesothelial cells
Abstract
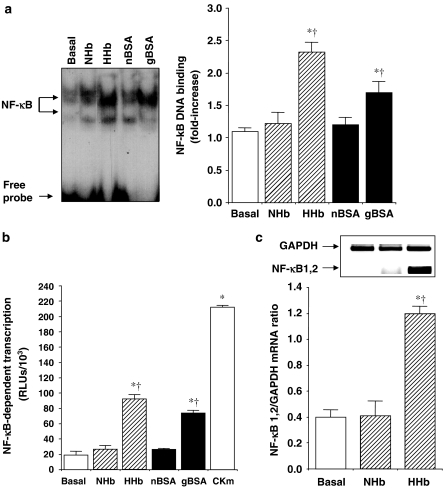
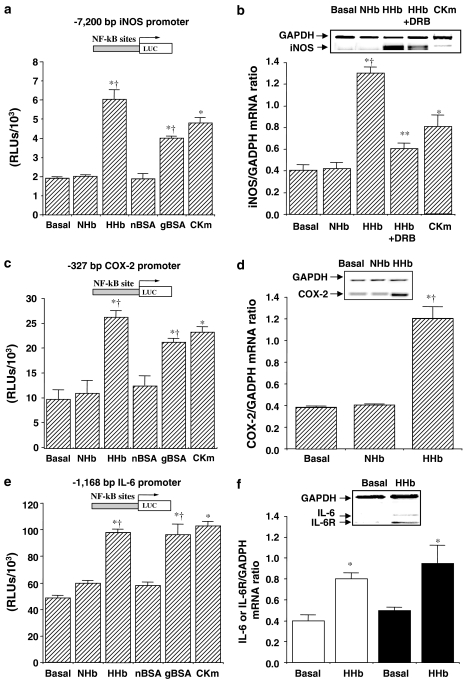
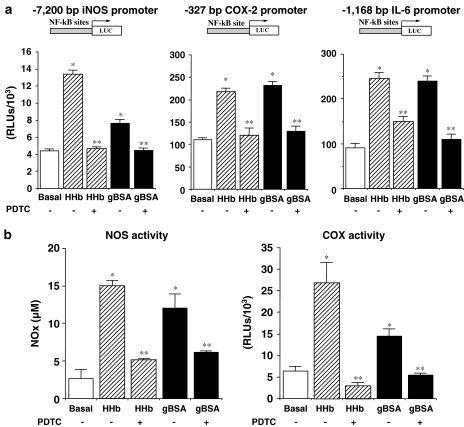
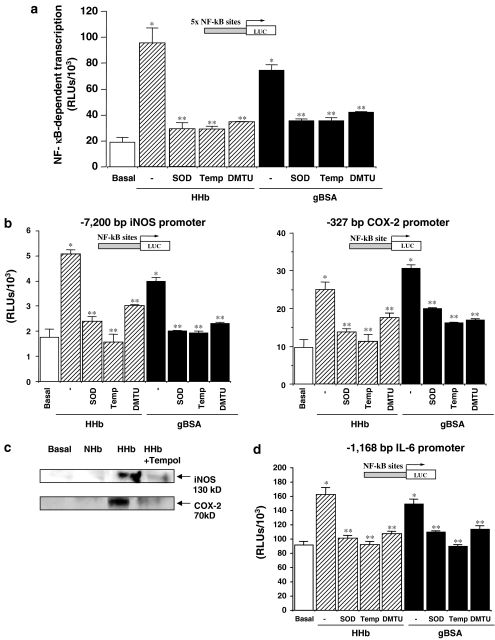
Diabetes mellitus leads to a high incidence of several so-called complications, sharing similar pathophysiological features in several territories. Previous reports points at early nonenzymatic glycosylation products (Amadori adducts) as mediators of diabetic vascular complications. In the present study, we analysed a possible role for Amadori adducts as stimulators of proinflammatory pathways in human peritoneal mesothelial cells (HPMCs). Cultured HPMCs isolated from 13 different patients (mean age 38.7+/-16 years) were exposed to different Amadori adducts, that is, highly glycated haemoglobin (10 nM) and glycated bovine serum albumin (0.25 mg ml(-1)), as well as to their respective low glycosylation controls. Amadori adducts, but not their respective controls, elicited a marked increase of NF-kappaB activation, as determined by electromobility shift assays and transient transfection experiments. Additionally, Amadori adducts significantly increased the production of NF-kappaB-related proinflammatory molecules, including cytokines, such as TNF-alpha, IL-1beta or IL-6, and enzymes, such as cyclooxygenase-2 and inducible nitric oxide (NO) synthase, this latter leading to the release of NO by HPMCs. The effects of Amadori adducts were mediated by different reactive oxygen and nitrosative species (e.g. superoxide anions, hydroxyl radicals, and peroxynitrite), as they were blunted by coincubation with the appropriate scavengers. Furthermore, NO generated upon exposure to Amadori adducts further stimulated NF-kappaB activation, either directly or after combination with superoxide anions to form peroxynitrite. We conclude that Amadori adducts can favour peritoneal inflammation by exacerbating changes in NO synthesis pathway and triggering NF-kappaB-related proinflammatory signals in human mesothelial cells.
Figures
Similar articles
-
Pro-inflammatory effects of early non-enzymatic glycated proteins in human mesothelial cells vary with cell donor's age.Br J Pharmacol. 2006 Dec;149(8):979-87. doi: 10.1038/sj.bjp.0706864. Epub 2006 Oct 30. Br J Pharmacol. 2006. PMID: 17075573 Free PMC article.
-
Glycosylated human oxyhaemoglobin activates nuclear factor-kappaB and activator protein-1 in cultured human aortic smooth muscle.Br J Pharmacol. 2003 Oct;140(4):681-90. doi: 10.1038/sj.bjp.0705483. Epub 2003 Sep 22. Br J Pharmacol. 2003. PMID: 14504138 Free PMC article.
-
Polyphenols Attenuate Highly-Glycosylated Haemoglobin-Induced Damage in Human Peritoneal Mesothelial Cells.Antioxidants (Basel). 2020 Jul 1;9(7):572. doi: 10.3390/antiox9070572. Antioxidants (Basel). 2020. PMID: 32630324 Free PMC article.
-
Pathophysiological role of Amadori-glycated proteins in diabetic microangiopathy.Semin Vasc Med. 2002 May;2(2):191-7. doi: 10.1055/s-2002-32042. Semin Vasc Med. 2002. PMID: 16222610 Review.
-
Amadori albumin in diabetic nephropathy.Indian J Endocrinol Metab. 2015 Jan-Feb;19(1):39-46. doi: 10.4103/2230-8210.146863. Indian J Endocrinol Metab. 2015. PMID: 25593824 Free PMC article. Review.
Cited by
-
Advanced Glycation End-Products Acting as Immunomodulators for Chronic Inflammation, Inflammaging and Carcinogenesis in Patients with Diabetes and Immune-Related Diseases.Biomedicines. 2024 Jul 31;12(8):1699. doi: 10.3390/biomedicines12081699. Biomedicines. 2024. PMID: 39200164 Free PMC article. Review.
-
Glycated albumin activates NADPH oxidase in rat mesangial cells through up-regulation of p47phox.Biochem Biophys Res Commun. 2010 Jun 18;397(1):5-11. doi: 10.1016/j.bbrc.2010.04.084. Epub 2010 Apr 23. Biochem Biophys Res Commun. 2010. PMID: 20399741 Free PMC article.
-
The Beneficial Effects of Actinomycetales Immune Modulators in the Pancreas of Diabetic Rats.Adv Pharm Bull. 2021 Feb;11(2):371-377. doi: 10.34172/apb.2021.035. Epub 2020 Apr 22. Adv Pharm Bull. 2021. PMID: 33880360 Free PMC article.
-
Paricalcitol reduces peritoneal fibrosis in mice through the activation of regulatory T cells and reduction in IL-17 production.PLoS One. 2014 Oct 3;9(10):e108477. doi: 10.1371/journal.pone.0108477. eCollection 2014. PLoS One. 2014. PMID: 25279459 Free PMC article.
-
Inhibition of NF-kappaB with Dehydroxymethylepoxyquinomicin modifies the function of human peritoneal mesothelial cells.Am J Transl Res. 2016 Dec 15;8(12):5756-5765. eCollection 2016. Am J Transl Res. 2016. PMID: 28078047 Free PMC article.
References
-
- AMORE A., CIRINA P., MITOLA S., PERUZZI L., GIANOGLIO B., RABBONE I., SACCHETTI C., CERUTTI F., GRILLO C., COPPO R. Nonenzymatically glycated albumin (Amadori adducts) enhances nitric oxide synthase activity and gene expression in endothelial cells. Kidney Int. 1997;51:27–35. - PubMed
-
- ANGULO J., SÁNCHEZ-FERRER C.F., PEIRÓ C., MARÍN J., RODRÍGUEZ-MAÑAS L. Impairment of endothelium-dependent relaxations by increasing percentages of glycated human haemoglobin. Hypertension. 1996;28:583–592. - PubMed
-
- BERGHE W.V., PLAISANCE S., BOONE E., DE BOSSCHER K., SCHIMITZ M.L., FIERS W., HAEGEMAN G. P38 and extracellular signal-regulated kinase mitogen-activated protein kinase pathway are regulated for nuclear-factor κB p65 transactivation mediated by tumor necrosis factor. J. Biol. Chem. 1998;273:3285–3290. - PubMed
-
- BODEGA F., ZACCHI L., AGOSTINI E. Albumin transcytosis in mesothelium. Am. J. Physiol. Lung Cell. Mol. Physiol. 2002;282:L3–L11. - PubMed
-
- BOULANGER E., WAUTIER M.P., WAUTIER J.C., BOVAL B., PANIS Y., WERNERT N., DANZE P.M., DEQUIEDT P. AGEs bind to mesothelial cells via RAGE and stimulate VCAM-1 expression. Kidney Int. 2002;61:148–156. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
