

Defining electrical communication in skeletal muscle resistance arteries: a computational approach
- PMID: 16002449
- PMCID: PMC1474767
- DOI: 10.1113/jphysiol.2005.090233
Defining electrical communication in skeletal muscle resistance arteries: a computational approach
Abstract
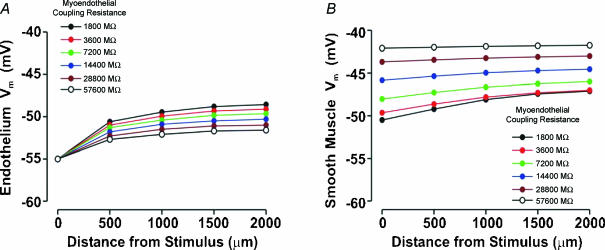
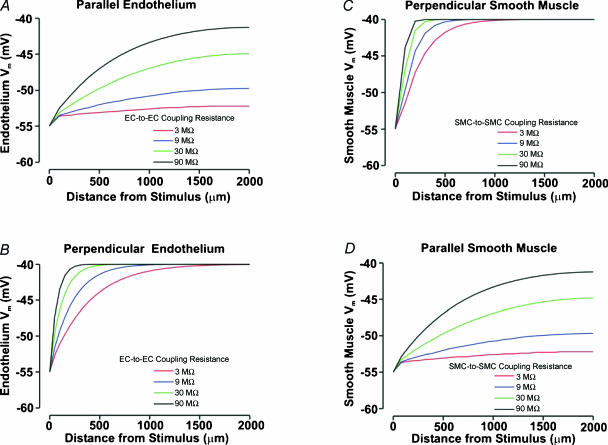
Vascular cells communicate electrically to coordinate their activity and control tissue blood flow. To foster a quantitative understanding of this fundamental process, we developed a computational model that was structured to mimic a skeletal muscle resistance artery. Each endothelial cell and smooth muscle cell in our virtual artery was treated as the electrical equivalent of a capacitor coupled in parallel with a non-linear resistor representing ionic conductance; intercellular gap junctions were represented by ohmic resistors. Simulations revealed that the vessel wall is not a syncytium in which electrical stimuli spread equally to all constitutive cells. Indeed, electrical signals spread in a differential manner among and between endothelial cells and smooth muscle cells according to the initial stimulus. The predictions of our model agree with physiological data from the feed artery of the hamster retractor muscle. Cell orientation and coupling resistance were the principal factors that enable electrical signals to spread differentially along and between the two cell types. Our computational observations also illustrated how gap junctional coupling enables the vessel wall to filter and transform transient electrical events into sustained voltage responses. Functionally, differential electrical communication would permit discrete regions of smooth muscle activity to locally regulate blood flow and the endothelium to coordinate regional changes in tissue perfusion.
Figures







References
-
- Bartlett IS, Segal SS. Resolution of smooth muscle and endothelial pathways for conduction along hamster cheek pouch arterioles. Am J Physiol. 2000;278:H604–H612. - PubMed
-
- Crane GJ, Hines ML, Neild TO. Simulating the spread of membrane potential changes in arteriolar networks. Microcirculation. 2001;8:33–43. - PubMed
-
- de Wit C, Roos F, Bolz SS, Kirchhoff S, Kruger O, Willecke K, Pohl U. Impaired conduction of vasodilation along arterioles in connexin40-deficient mice. Circ Res. 2000;86:649–655. - PubMed
-
- Emerson GG, Neild TO, Segal SS. Conduction of hyperpolarization along hamster feed arteries: augmentation by acetylcholine. Am J Physiol. 2002;283:H102–H109. - PubMed
-
- Emerson GG, Segal SS. Endothelial cell pathway for conduction of hyperpolarization and vasodilation along hamster feed artery. Circ Res. 2000a;86:94–100. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous