

Kinetics of the reverse mode of the Na+/glucose cotransporter
- PMID: 16007500
- PMCID: PMC3000923
- DOI: 10.1007/s00232-005-0743-x
Kinetics of the reverse mode of the Na+/glucose cotransporter
Abstract
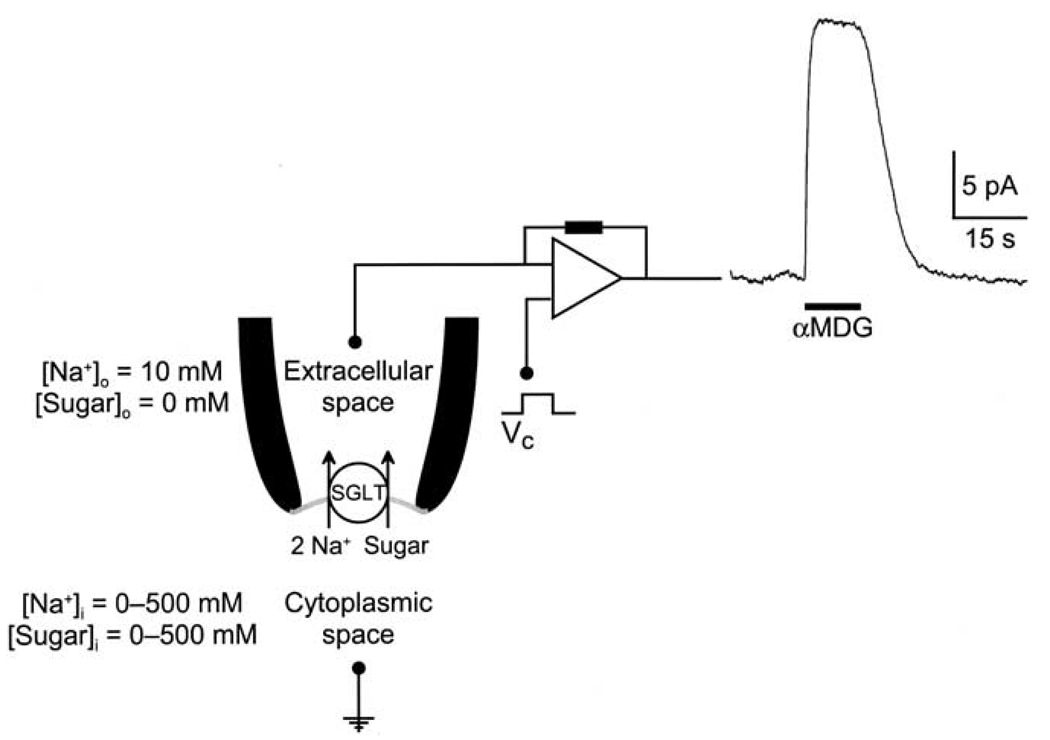
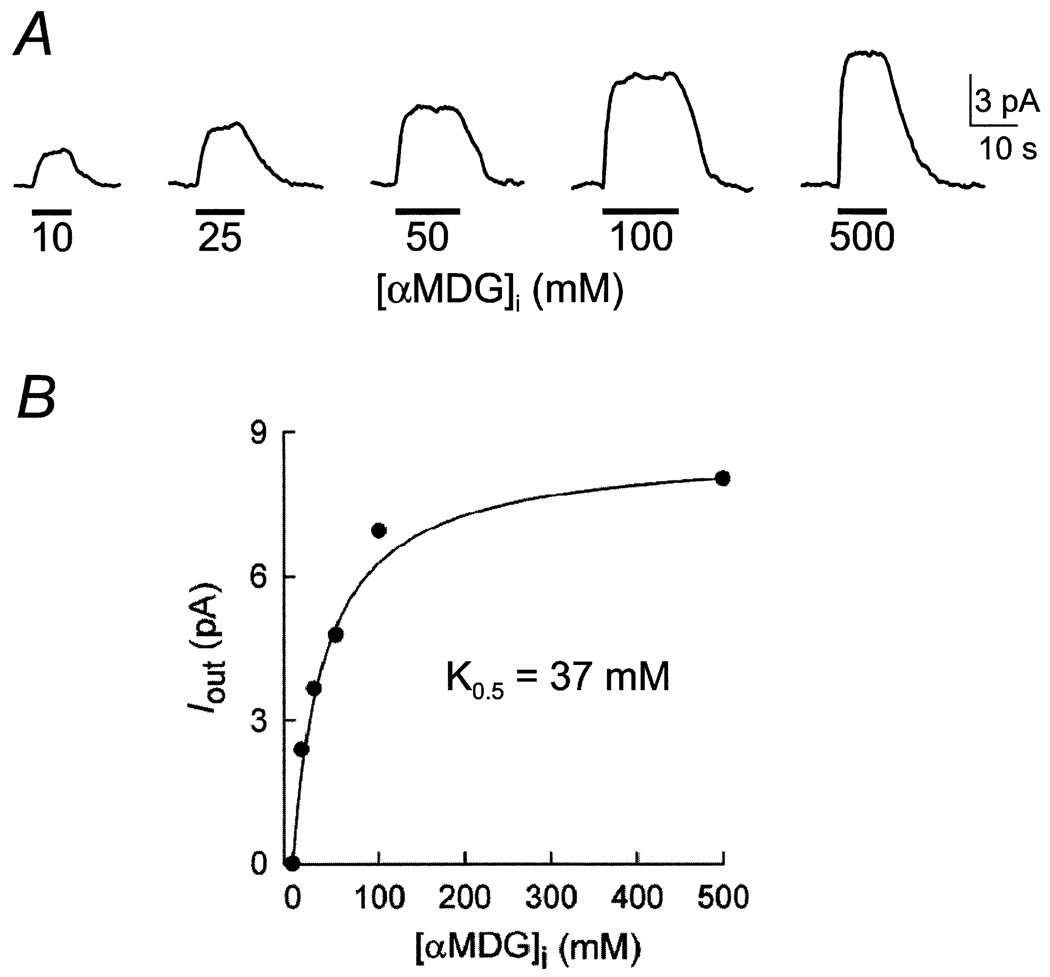
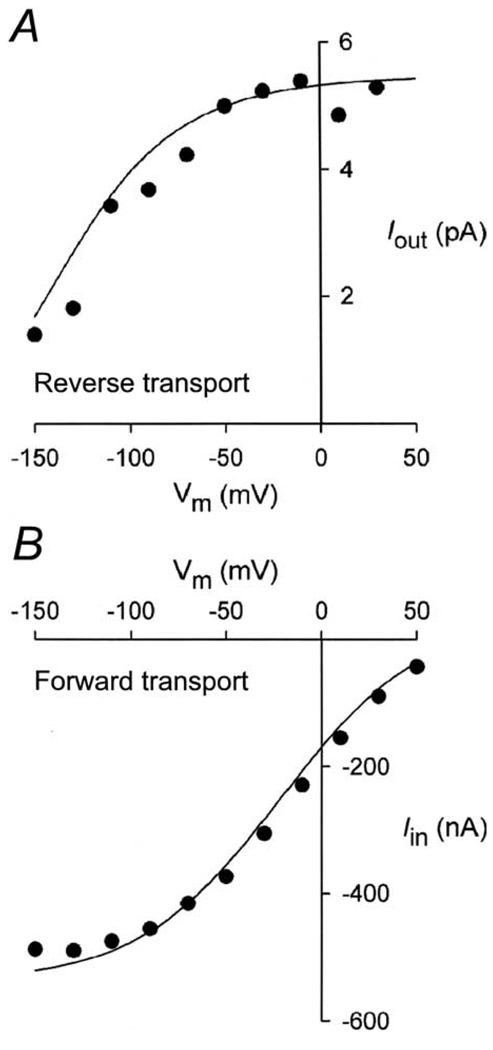
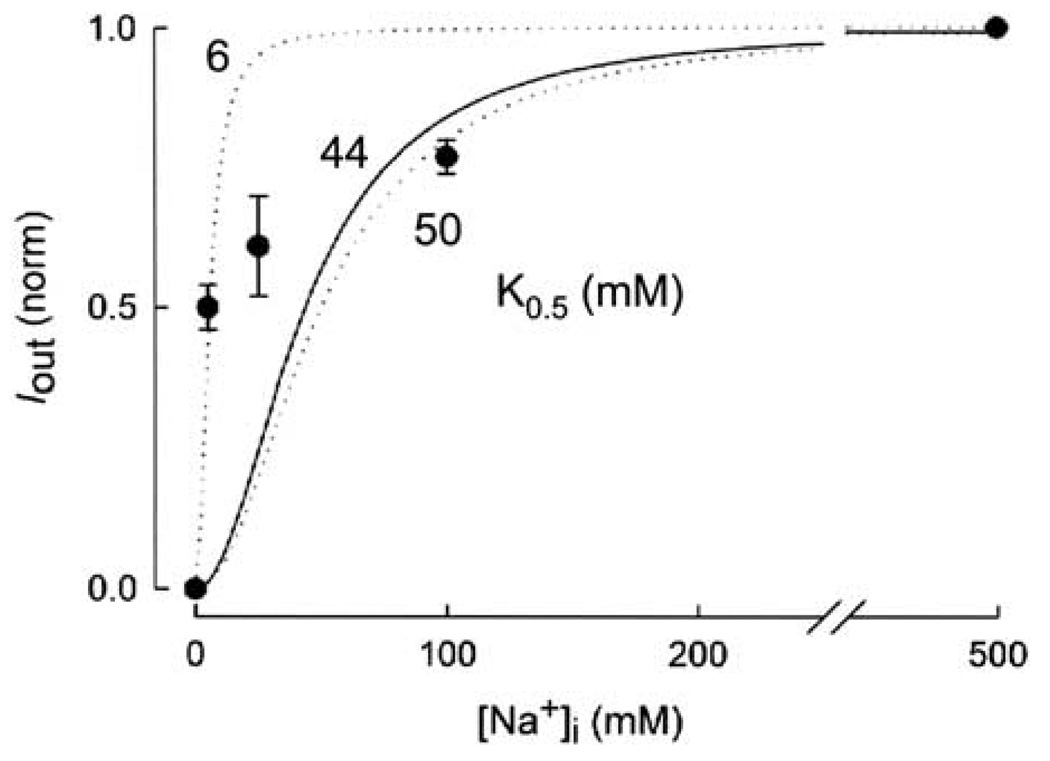
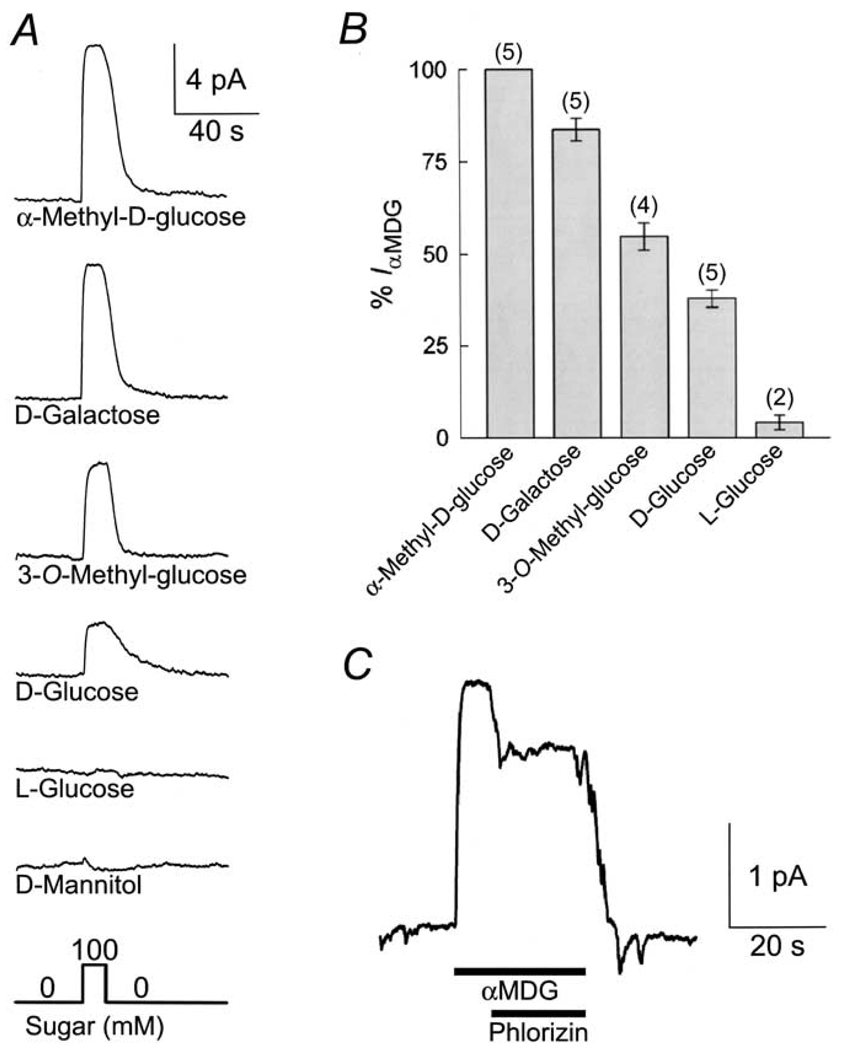
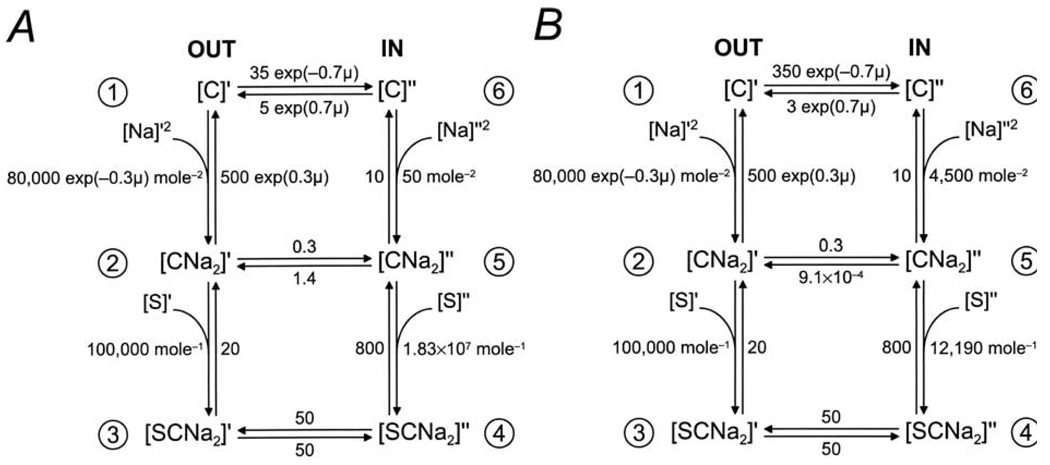
This study investigates the reverse mode of the Na(+)/glucose cotransporter (SGLT1). In giant excised inside-out membrane patches from Xenopus laevis oocytes expressing rabbit SGLT1, application of alpha-methyl-D: -glucopyranoside (alphaMDG) to the cytoplasmic solution induced an outward current from cytosolic to external membrane surface. The outward current was Na(+)- and sugar-dependent, and was blocked by phlorizin, a specific inhibitor of SGLT1. The current-voltage relationship saturated at positive membrane voltages (30-50 mV), and approached zero at -150 mV. The half-maximal concentration for alphaMDG-evoked outward current (K(0.5) (alphaMDG)) was 35 mM (at 0 mV). In comparison, K(0.5) (alphaMDG) for forward sugar transport was 0.15 mM (at 0 mV). K(0.5) (Na) was similar for forward and reverse transport ( approximately 35 mM at 0 mV). Specificity of SGLT1 for reverse transport was: alphaMDG (1.0) > D: -galactose (0.84) > 3-O-methyl-glucose (0.55) > D: -glucose (0.38), whereas for forward transport, specificity was: alphaMDG approximately D: -glucose approximately D: -galactose > 3-O-methyl-glucose. Thus there is an asymmetry in sugar kinetics and specificity between forward and reverse modes. Computer simulations showed that a 6-state kinetic model for SGLT1 can account for Na(+)/sugar cotransport and its voltage dependence in both the forward and reverse modes at saturating sodium concentrations. Our data indicate that under physiological conditions, the transporter is poised to accumulate sugar efficiently in the enterocyte.
Figures
References
-
- Abramson J, Smirnova I, Kasho V, Verner G, Kaback HR, Iwata S. Structure and mechanism of the lactose permease of Escherichia coli. Science. 2003;301:610–615. - PubMed
-
- Birnir B, Loo DDF, Wright EM. Voltage-clamp studies of the Na+/glucose cotransporter cloned from rabbit small intestine. Pfluegers Arch. 1991;418:79–85. - PubMed
-
- Díez-Sampedro A, Wright EM, Hirayama BA. Residue 457 controls sugar binding and transport in the Na+/glucose cotransporter. J. Biol. Chem. 2001;276:49188–48194. - PubMed
-
- Eskandari S, Loo DDF, Wright EM. Functional asymmetry of the sodium/glucose cotransported. FASEB J. Abs. 1999:399.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
