

Aquaporin 7 deficiency is associated with development of obesity through activation of adipose glycerol kinase
- PMID: 16009937
- PMCID: PMC1182435
- DOI: 10.1073/pnas.0503291102
Aquaporin 7 deficiency is associated with development of obesity through activation of adipose glycerol kinase
Abstract
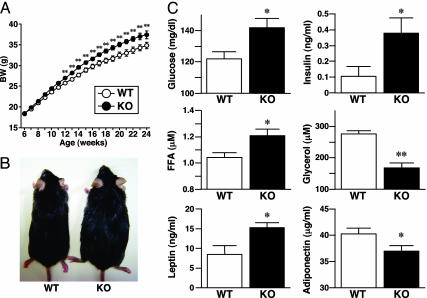
In adipocytes, hydrolysis of triglycerides results in the release of free fatty acids and glycerol. Aquaporin 7 (AQP7), a member of aquaglyceroporins, is known to permeabilize glycerol and water. We recently generated Aqp7-knockout (KO) mice and demonstrated that such mice have low plasma glycerol levels and impaired glycerol release in response to beta3-adrenergic agonist, suggesting that AQP7 acts as a glycerol gateway molecule in adipocytes for the efficient release of glycerol in vivo. Although there was no difference in body weight between WT and KO mice until 10 weeks of age, here we found that KO mice developed adult-onset obesity. The body weight and fat mass increased significantly in KO mice compared with WT mice after 12 weeks of age. Adipocytes of KO mice were large and exhibited accumulation of triglycerides compared with WT mice. The KO mice developed obesity and insulin resistance even at a young age after consumption of high-fat/high-sucrose diet. We demonstrated the enhanced glycerol kinase enzymatic activity in Aqp7-KO and -knockdown adipocytes. A series of our results indicate that AQP7 disruption elevates adipose glycerol kinase activity, accelerates triglycerides synthesis in adipocytes, and, finally, develops obesity.
Figures





Comment in
-
Obesity and metabolic perturbations after loss of aquaporin 7, the adipose glycerol transporter.Proc Natl Acad Sci U S A. 2005 Aug 2;102(31):10759-60. doi: 10.1073/pnas.0504965102. Epub 2005 Jul 26. Proc Natl Acad Sci U S A. 2005. PMID: 16046537 Free PMC article. No abstract available.
References
-
- Denker, B. M., Smith, B. L., Kuhajda, F. P. & Agre, P. (1988) J. Biol. Chem. 263, 15634-15642. - PubMed
-
- Agre, P. & Kozono, D. (2003) FEBS Lett. 555, 72-78. - PubMed
-
- Preston, G. M., Carroll, T. P., Guggino, W. B. & Agre, P. (1992) Science 256, 385-387. - PubMed
-
- Giles, J. (2003) Nature 425, 651. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
