

Engineering cytoplasmic male sterility via the chloroplast genome by expression of {beta}-ketothiolase
- PMID: 16009998
- PMCID: PMC1176397
- DOI: 10.1104/pp.104.057729
Engineering cytoplasmic male sterility via the chloroplast genome by expression of {beta}-ketothiolase
Abstract
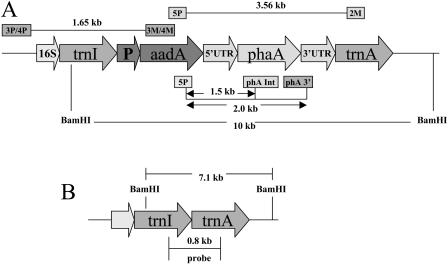
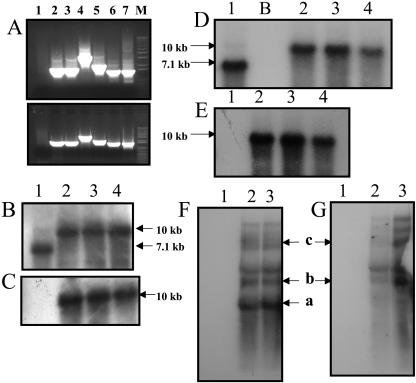
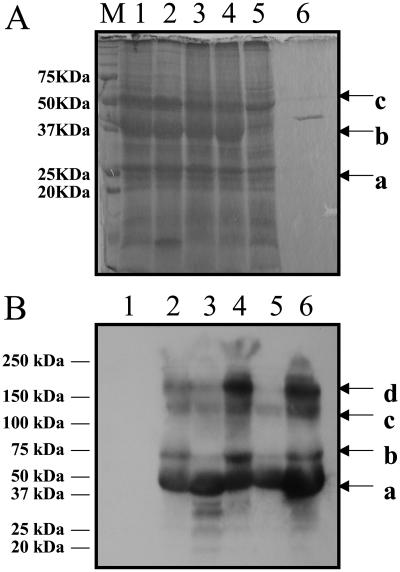
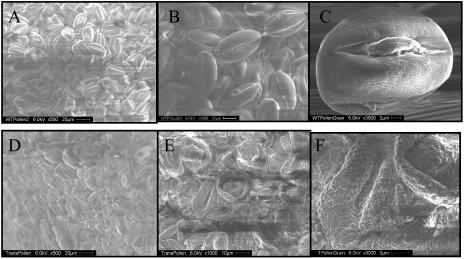
While investigating expression of the polydroxybutyrate pathway in transgenic chloroplasts, we addressed the specific role of beta-ketothiolase. Therefore, we expressed the phaA gene via the chloroplast genome. Prior attempts to express the phaA gene in transgenic plants were unsuccessful. We studied the effect of light regulation of the phaA gene using the psbA promoter and 5' untranslated region, and evaluated expression under different photoperiods. Stable transgene integration into the chloroplast genome and homoplasmy were confirmed by Southern analysis. The phaA gene was efficiently transcribed in all tissue types examined, including leaves, flowers, and anthers. Coomassie-stained gel and western blots confirmed hyperexpression of beta-ketothiolase in leaves and anthers, with proportionately high levels of enzyme activity. The transgenic lines were normal except for the male-sterile phenotype, lacking pollen. Scanning electron microscopy revealed a collapsed morphology of the pollen grains. Floral developmental studies revealed that transgenic lines showed an accelerated pattern of anther development, affecting their maturation, and resulted in aberrant tissue patterns. Abnormal thickening of the outer wall, enlarged endothecium, and vacuolation affected pollen grains and resulted in the irregular shape or collapsed phenotype. Reversibility of the male-sterile phenotype was observed under continuous illumination, resulting in viable pollen and copious amount of seeds. This study results in the first engineered cytoplasmic male-sterility system in plants, offers a new tool for transgene containment for both nuclear and organelle genomes, and provides an expedient mechanism for F(1) hybrid seed production.
Figures


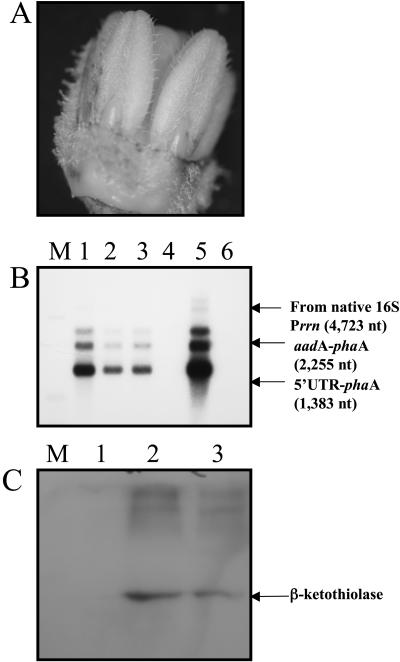
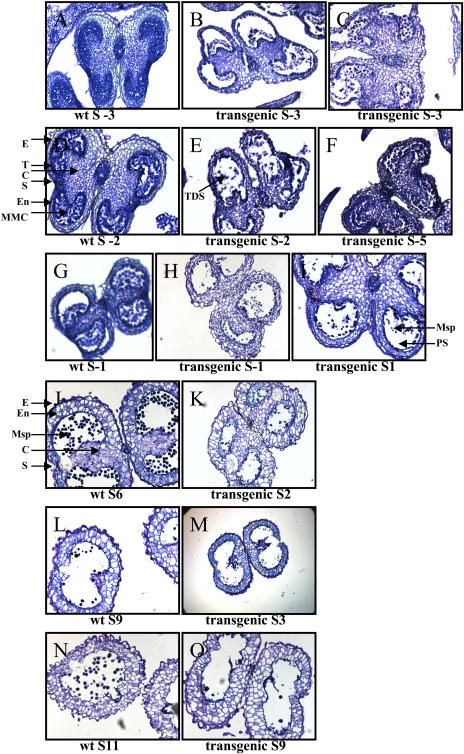

References
-
- Ariizumi T, Hatakeyama K, Hinata K, Inatsugi R, Nishida I, Sato S, Kato T, Tabata S, Toriyama K (2004) Disruption of the novel plant protein NEF1 affects lipid accumulation in the plastids of the tapetum and exine formation of pollen, resulting in male sterility in Arabidopsis thaliana. Plant J 39: 170–181 - PubMed
-
- Bateson W, Gairdner AE (1921) Male sterility in flax subject to two types of segregation. J Genet 11: 269–275
-
- Bohmert K, Balbo I, Kopka J, Mittendorf V, Nawrath C, Poirier Y, Tischendorf G, Trethewey RN, Willmitzer L (2000) Transgenic Arabidopsis plants can accumulate polyhydroxybutyrate to up to 4% of their fresh weight. Planta 211: 841–845 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
