

Utilization of homotypic and heterotypic proteins of vesicular stomatitis virus by defective interfering particle genomes for RNA replication and virion assembly: implications for the mechanism of homologous viral interference
- PMID: 16014921
- PMCID: PMC1181566
- DOI: 10.1128/JVI.79.15.9588-9596.2005
Utilization of homotypic and heterotypic proteins of vesicular stomatitis virus by defective interfering particle genomes for RNA replication and virion assembly: implications for the mechanism of homologous viral interference
Abstract
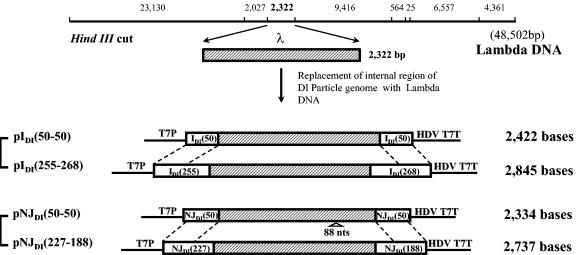
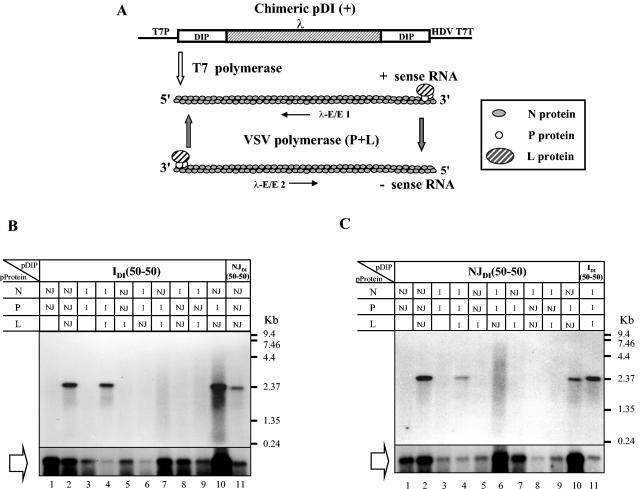
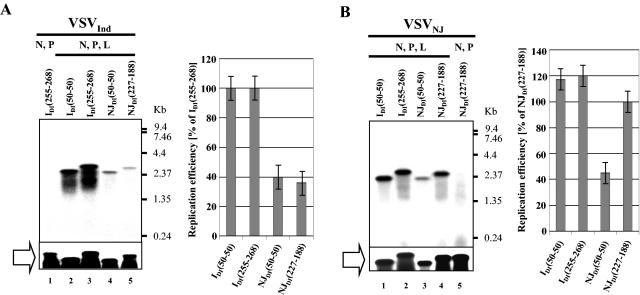
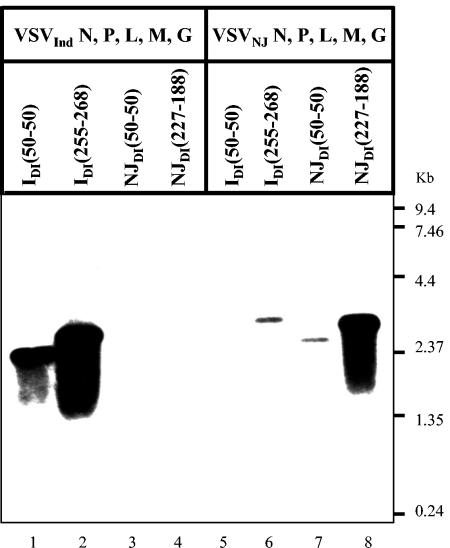
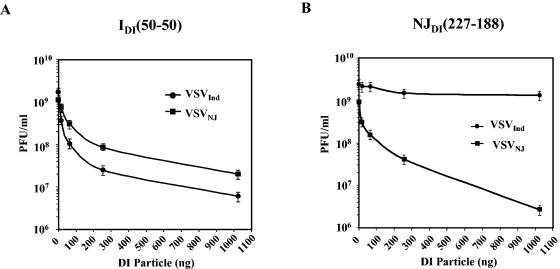
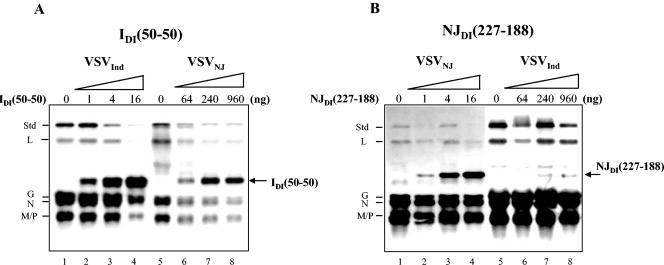
Defective interfering (DI) particles of Indiana serotype of vesicular stomatitis virus (VSV(Ind)) are capable of interfering with the replication of both homotypic VSV(Ind) and heterotypic New Jersey serotype (VSV(NJ)) standard virus. In contrast, DI particles from VSV(NJ) do not interfere with the replication of VSV(Ind) standard virus but do interfere with VSV(NJ) replication. The differences in the interfering activities of VSV(Ind) DI particles and VSV(NJ) DI particles against heterotypic standard virus were investigated. We examined the utilization of homotypic and heterotypic VSV proteins by DI particle genomic RNAs for replication and maturation into infectious DI particles. Here we show that the RNA-nucleocapsid protein (N) complex of one serotype does not utilize the polymerase complex (P and L) of the other serotype for RNA synthesis, while DI particle genomic RNAs of both serotypes can utilize the N, P, and L proteins of either serotype without serotypic restriction but with differing efficiencies as long as all three proteins are derived from the same serotype. The genomic RNAs of VSV(Ind) DI particles assembled and matured into DI particles by using either homotypic or heterotypic viral proteins. In contrast, VSV(NJ) DI particles could assemble only with homotypic VSV(NJ) viral proteins, although the genomic RNAs of VSV(NJ) DI particles could be replicated by using heterotypic VSV(Ind) N, P, and L proteins. Thus, we concluded that both efficient RNA replication and assembly of DI particles are required for the heterotypic interference by VSV DI particles.
Figures
Similar articles
-
Characterization of virulent isolates of vesicular stomatitis virus in relation to interference by defective particles.Microb Pathog. 1986 Apr;1(2):205-15. doi: 10.1016/0882-4010(86)90022-7. Microb Pathog. 1986. PMID: 2854597
-
Determinants of serotype specificity in transcription of vesicular stomatitis virus synthetic nucleocapsids.Virology. 1994 Feb 15;199(1):11-9. doi: 10.1006/viro.1994.1093. Virology. 1994. PMID: 8116233
-
Cells that express all five proteins of vesicular stomatitis virus from cloned cDNAs support replication, assembly, and budding of defective interfering particles.Proc Natl Acad Sci U S A. 1991 Feb 15;88(4):1379-83. doi: 10.1073/pnas.88.4.1379. Proc Natl Acad Sci U S A. 1991. PMID: 1847519 Free PMC article.
-
Terminal sequences of vesicular stomatitis virus RNA are both complementary and conserved.J Virol. 1979 Oct;32(1):167-74. doi: 10.1128/JVI.32.1.167-174.1979. J Virol. 1979. PMID: 232169 Free PMC article.
-
Defective interfering viruses and their potential as antiviral agents.Rev Med Virol. 2010 Jan;20(1):51-62. doi: 10.1002/rmv.641. Rev Med Virol. 2010. PMID: 20041441 Review.
Cited by
-
Asymmetric competitive suppression between strains of dengue virus.BMC Microbiol. 2008 Feb 8;8:28. doi: 10.1186/1471-2180-8-28. BMC Microbiol. 2008. PMID: 18261207 Free PMC article.
-
Limited interference at the early stage of infection between two recombinant novirhabdoviruses: viral hemorrhagic septicemia virus and infectious hematopoietic necrosis virus.J Virol. 2010 Oct;84(19):10038-50. doi: 10.1128/JVI.00343-10. Epub 2010 Jul 14. J Virol. 2010. PMID: 20631140 Free PMC article.
-
Viral Coinfections.Viruses. 2022 Nov 26;14(12):2645. doi: 10.3390/v14122645. Viruses. 2022. PMID: 36560647 Free PMC article. Review.
-
Creation of matrix protein gene variants of two serotypes of vesicular stomatitis virus as prime-boost vaccine vectors.J Virol. 2015 Jun;89(12):6338-51. doi: 10.1128/JVI.00222-15. Epub 2015 Apr 8. J Virol. 2015. PMID: 25855732 Free PMC article.
-
Virological and Immunological Outcomes of Coinfections.Clin Microbiol Rev. 2018 Jul 5;31(4):e00111-17. doi: 10.1128/CMR.00111-17. Print 2018 Oct. Clin Microbiol Rev. 2018. PMID: 29976554 Free PMC article. Review.
References
-
- Bishop, D. H., S. U. Emerson, and A. Flamand. 1974. Reconstitution of infectivity and transcriptase activity of homologous and heterologous viruses: vesicular stomatitis (Indiana serotype), Chandipura, vesicular stomatitis (New Jersey serotype), and Cocal viruses. J. Virol. 14:139-144. - PMC - PubMed
-
- Brown, T., K. Mackey, and T. Du. 1997. Analysis of RNA by Northern and slot blot hybridization, p. 4.9.1-4.9.16. In R. B. Frederick, M. Ausubel, R. E. Kingston, D. D. Moore, J. G. Seidman, J. A. Smith, and K. Struhl (ed.), Current protocols in molecular biology. John Wiley and Sons, New York, N.Y. - PubMed
-
- Choi, W. Y. 1997. Molecular biological characterization of defective interfering particles of vesicular stomatitis virus. University of Ottawa, Ottawa, Canada.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
