

mRNA decay during herpes simplex virus (HSV) infections: protein-protein interactions involving the HSV virion host shutoff protein and translation factors eIF4H and eIF4A
- PMID: 16014927
- PMCID: PMC1181552
- DOI: 10.1128/JVI.79.15.9651-9664.2005
mRNA decay during herpes simplex virus (HSV) infections: protein-protein interactions involving the HSV virion host shutoff protein and translation factors eIF4H and eIF4A
Abstract
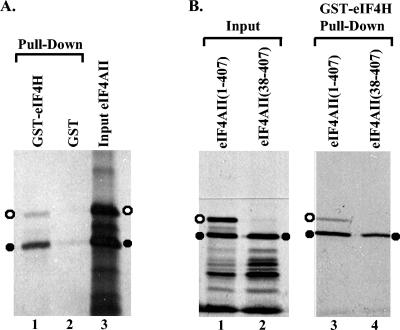
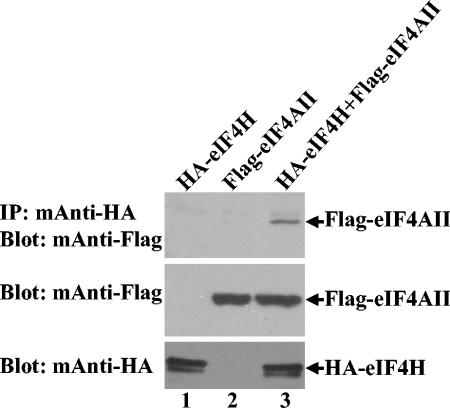
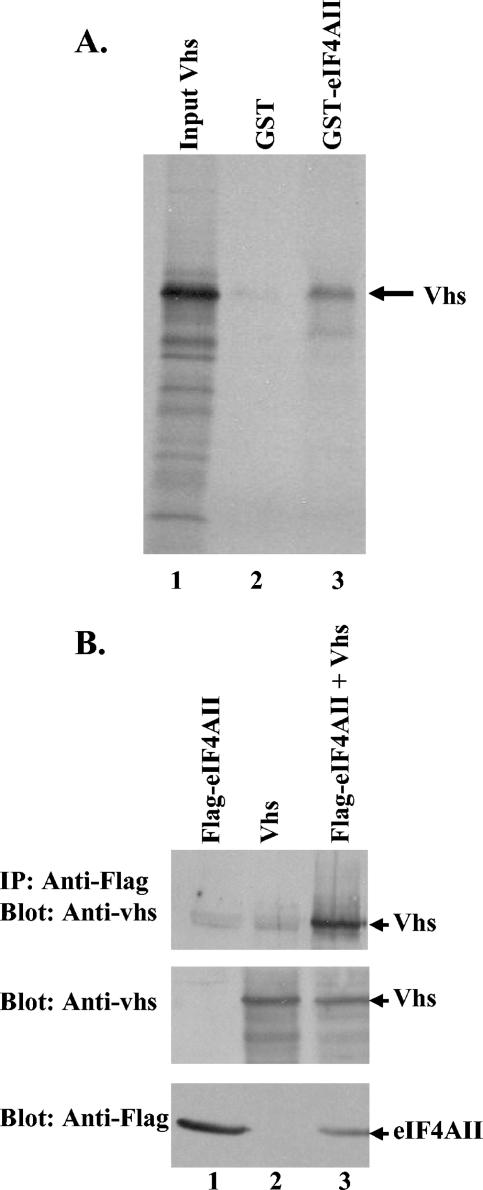
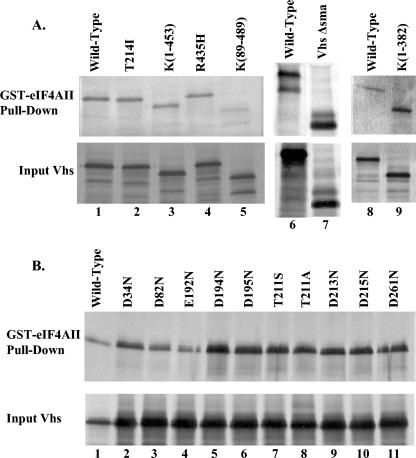
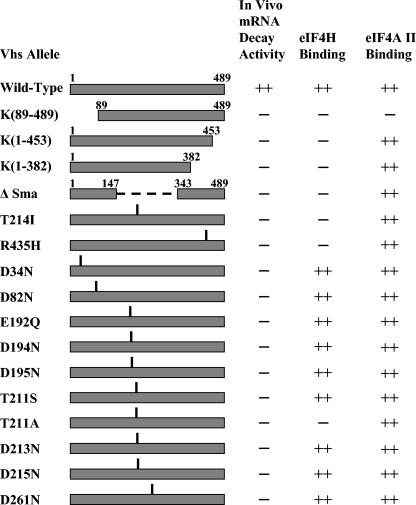
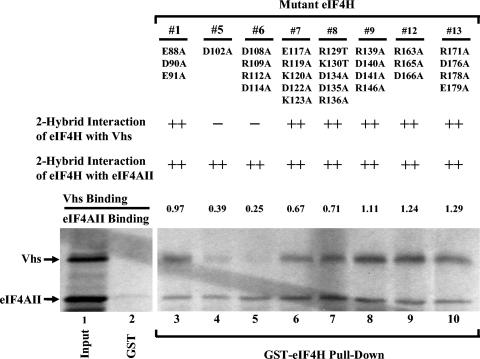
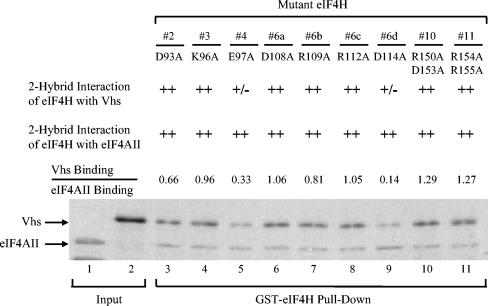
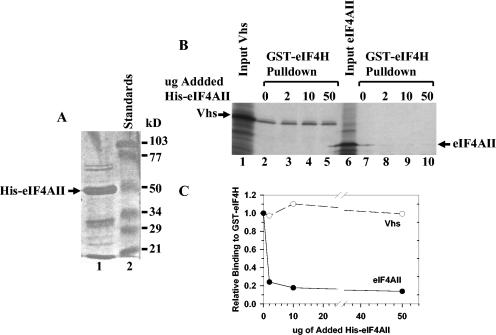
During lytic infections, the virion host shutoff (Vhs) protein of herpes simplex virus accelerates the degradation of both host and viral mRNAs. In so doing, it helps redirect the cell from host to viral protein synthesis and facilitates the sequential expression of different viral genes. Vhs interacts with the cellular translation initiation factor eIF4H, and several point mutations that abolish its mRNA degradative activity also abrogate its ability to bind eIF4H. In addition, a complex containing bacterially expressed Vhs and a glutathione S-transferase (GST)-eIF4H fusion protein has RNase activity. eIF4H shares a region of sequence homology with eIF4B, and it appears to be functionally similar in that both stimulate the RNA helicase activity of eIF4A, a component of the mRNA cap-binding complex eIF4F. We show that eIF4H interacts physically with eIF4A in the yeast two-hybrid system and in GST pull-down assays and that the two proteins can be coimmunoprecipitated from mammalian cells. Vhs also interacts with eIF4A in GST pull-down and coimmunoprecipitation assays. Site-directed mutagenesis of Vhs and eIF4H revealed residues of each that are important for their mutual interaction, but not for their interaction with eIF4A. Thus, Vhs, eIF4H, and eIF4A comprise a group of proteins, each of which is able to interact directly with the other two. Whether they interact simultaneously as a tripartite complex or sequentially is unclear. The data suggest a mechanism for linking the degradation of an mRNA to its translation and for targeting Vhs to mRNAs and to regions of translation initiation.
Figures

References
-
- Becker, Y., E. Tavor, Y. Asher, C. Berkowiltz, and M. Moyal. 1993. Effect of herpes simplex virus type-1 UL41 gene on the stability of mRNA from the cellular genes: beta-actin, fibronectin, glucose transporter-1, and docking protein, and on virus intraperitoneal pathogenicity of newborn mice. Virus Genes 7:133-143. - PubMed
-
- Callaghan, A. J., J. P. Aurikko, L. L. Ilag, G. J. Gunter, V. Chandran, K. Kuhnel, L. Poljak, A. J. Carpousis, C. V. Robinson, M. F. Symmons, and B. F. Luisi. 2004. Studies of the RNA degradosome-organizing domain of the Escherichia coli ribonuclease RNase E. J. Mol. Biol. 340:965-979. - PubMed
-
- Carpousis, A. J. 2002. The Escherichia coli RNA degradosome: structure, function and relationship in other ribonucleolytic multienzyme complexes. Biochem. Soc. Trans. 30:150-155. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
