

Mechanism of stimulation of plus-strand synthesis by an RNA replication enhancer in a tombusvirus
- PMID: 16014939
- PMCID: PMC1181556
- DOI: 10.1128/JVI.79.15.9777-9785.2005
Mechanism of stimulation of plus-strand synthesis by an RNA replication enhancer in a tombusvirus
Abstract
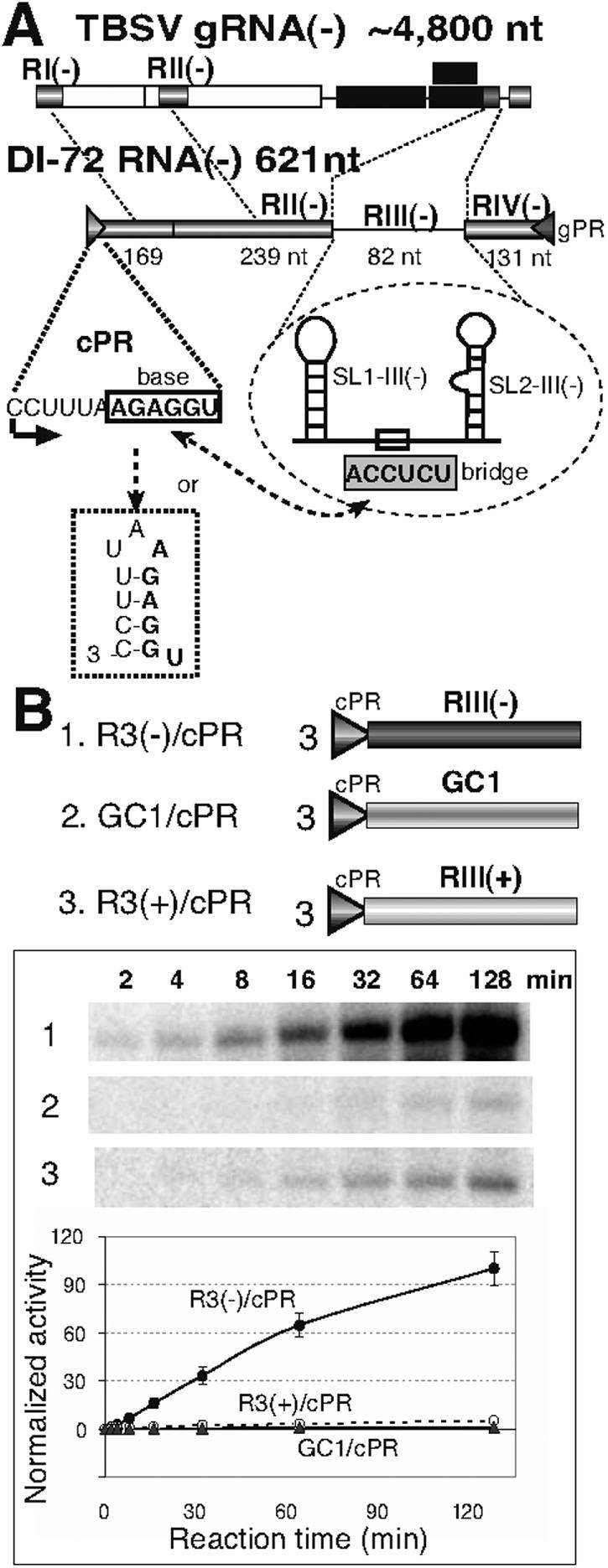
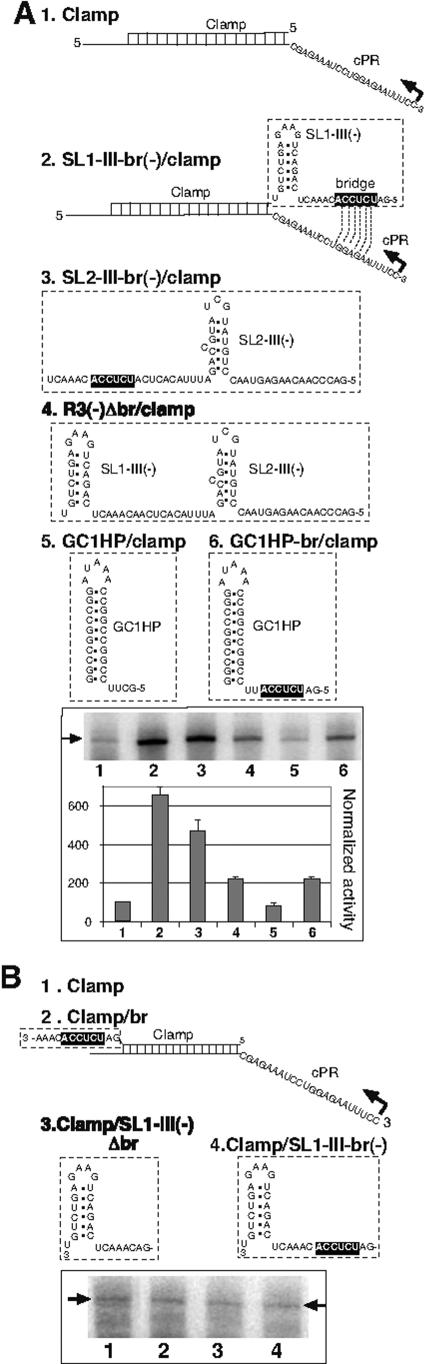
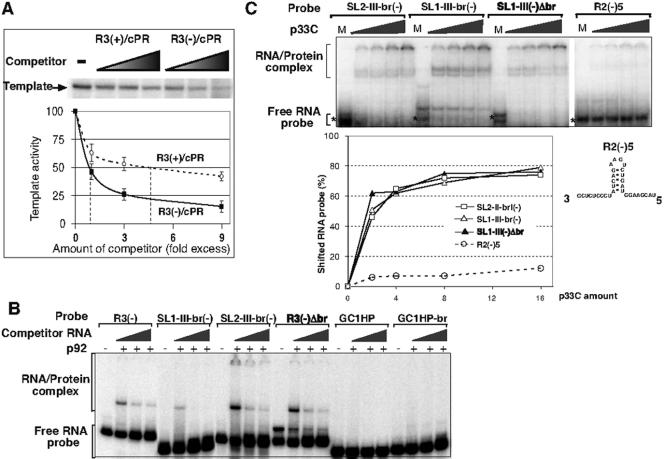
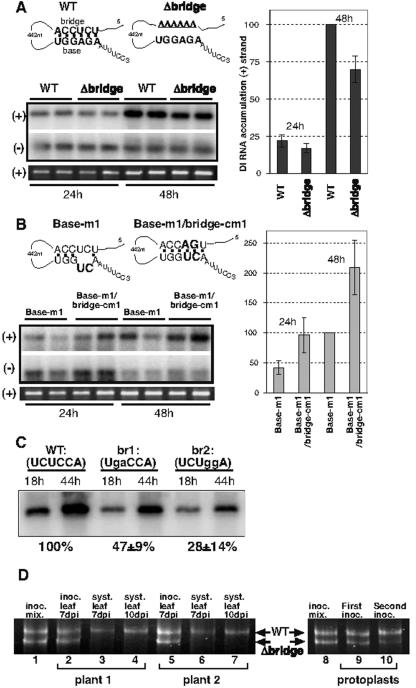
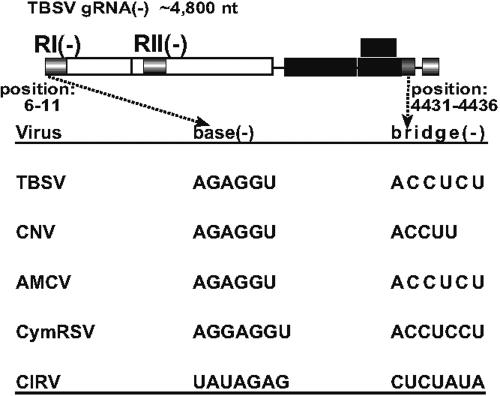
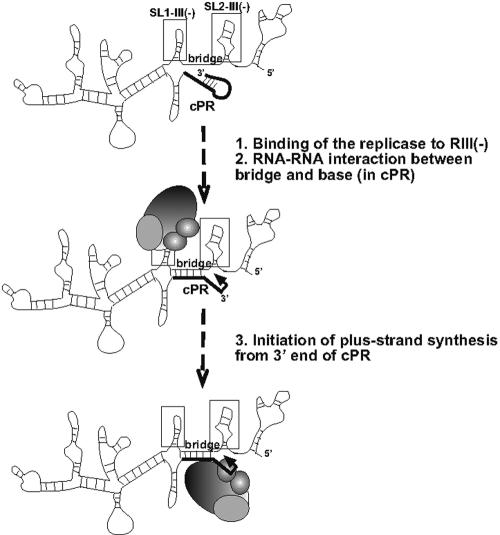
Replication of RNA viruses is regulated by cis-acting RNA elements, including promoters, replication silencers, and replication enhancers (REN). To dissect the function of an REN element involved in plus-strand RNA synthesis, we developed an in vitro trans-replication assay for tombusviruses, which are small plus-strand RNA viruses. In this assay, two RNA strands were tethered together via short complementary regions with the REN present in the nontemplate RNA, whereas the promoter was located in the template RNA. We found that the template activity of the tombusvirus replicase preparation was stimulated in trans by the REN, suggesting that the REN is a functional enhancer when located in the vicinity of the promoter. In addition, this study revealed that the REN has dual function during RNA synthesis. (i) It binds to the viral replicase. (ii) It interacts with the core plus-strand initiation promoter via a long-distance RNA-RNA interaction, which leads to stimulation of initiation of plus-strand RNA synthesis by the replicase in vitro. We also observed that this RNA-RNA interaction increased the in vivo accumulation and competitiveness of defective interfering RNA, a model template. We propose that REN is important for asymmetrical viral RNA replication that leads to more abundant plus-strand RNA progeny than the minus-strand intermediate, a hallmark of replication of plus-strand RNA viruses.
Figures
Similar articles
-
The RNA replication enhancer element of tombusviruses contains two interchangeable hairpins that are functional during plus-strand synthesis.J Virol. 2003 Jan;77(1):258-69. doi: 10.1128/jvi.77.1.258-269.2003. J Virol. 2003. PMID: 12477831 Free PMC article.
-
Heterologous RNA replication enhancer stimulates in vitro RNA synthesis and template-switching by the carmovirus, but not by the tombusvirus, RNA-dependent RNA polymerase: implication for modular evolution of RNA viruses.Virology. 2005 Oct 10;341(1):107-21. doi: 10.1016/j.virol.2005.06.042. Virology. 2005. PMID: 16083933
-
Enhancement of RNA synthesis by promoter duplication in tombusviruses.Virology. 2003 May 25;310(1):118-29. doi: 10.1016/s0042-6822(03)00105-3. Virology. 2003. PMID: 12788636
-
Yeast as a model host to dissect functions of viral and host factors in tombusvirus replication.Virology. 2006 Jan 5;344(1):211-20. doi: 10.1016/j.virol.2005.09.017. Virology. 2006. PMID: 16364751 Review.
-
Advances in the molecular biology of tombusviruses: gene expression, genome replication, and recombination.Prog Nucleic Acid Res Mol Biol. 2004;78:187-226. doi: 10.1016/S0079-6603(04)78005-8. Prog Nucleic Acid Res Mol Biol. 2004. PMID: 15210331 Review.
Cited by
-
Coordinated function of cellular DEAD-box helicases in suppression of viral RNA recombination and maintenance of viral genome integrity.PLoS Pathog. 2015 Feb 18;11(2):e1004680. doi: 10.1371/journal.ppat.1004680. eCollection 2015 Feb. PLoS Pathog. 2015. PMID: 25693185 Free PMC article.
-
RNA chaperone activity of the tombusviral p33 replication protein facilitates initiation of RNA synthesis by the viral RdRp in vitro.Virology. 2011 Jan 20;409(2):338-47. doi: 10.1016/j.virol.2010.10.015. Epub 2010 Nov 10. Virology. 2011. PMID: 21071052 Free PMC article.
-
A discontinuous RNA platform mediates RNA virus replication: building an integrated model for RNA-based regulation of viral processes.PLoS Pathog. 2009 Mar;5(3):e1000323. doi: 10.1371/journal.ppat.1000323. Epub 2009 Mar 6. PLoS Pathog. 2009. PMID: 19266082 Free PMC article.
-
RNA Structure Protects the 5' End of an Uncapped Tombusvirus RNA Genome from Xrn Digestion.J Virol. 2021 Sep 27;95(20):e0103421. doi: 10.1128/JVI.01034-21. Epub 2021 Aug 4. J Virol. 2021. PMID: 34346764 Free PMC article.
-
Host protein chaperones, RNA helicases and the ubiquitin network highlight the arms race for resources between tombusviruses and their hosts.Adv Virus Res. 2020;107:133-158. doi: 10.1016/bs.aivir.2020.06.006. Epub 2020 Jul 7. Adv Virus Res. 2020. PMID: 32711728 Free PMC article.
References
-
- Ahlquist, P. 2002. RNA-dependent RNA polymerases, viruses, and RNA silencing. Science 296:1270-1273. - PubMed
-
- Choi, I. R., M. Ostrovsky, G. Zhang, and K. A. White. 2001. Regulatory activity of distal and core RNA elements in Tombusvirus subgenomic mRNA2 transcription. J. Biol. Chem. 276:41761-41768. - PubMed
-
- Dreher, T. W. 1999. Functions of the 3′-untranslated regions of positive strand RNA viral genomes. Annu. Rev. Phytopathol. 37:151-174. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
