

Murine coronavirus requires lipid rafts for virus entry and cell-cell fusion but not for virus release
- PMID: 16014947
- PMCID: PMC1181594
- DOI: 10.1128/JVI.79.15.9862-9871.2005
Murine coronavirus requires lipid rafts for virus entry and cell-cell fusion but not for virus release
Abstract
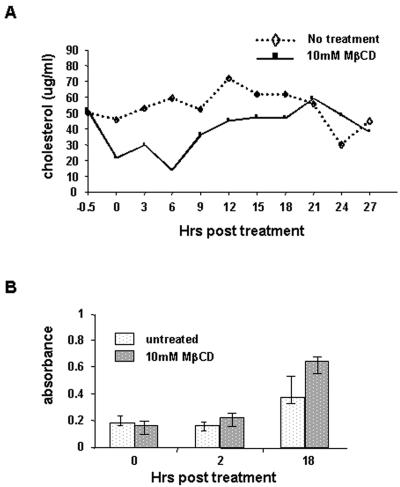
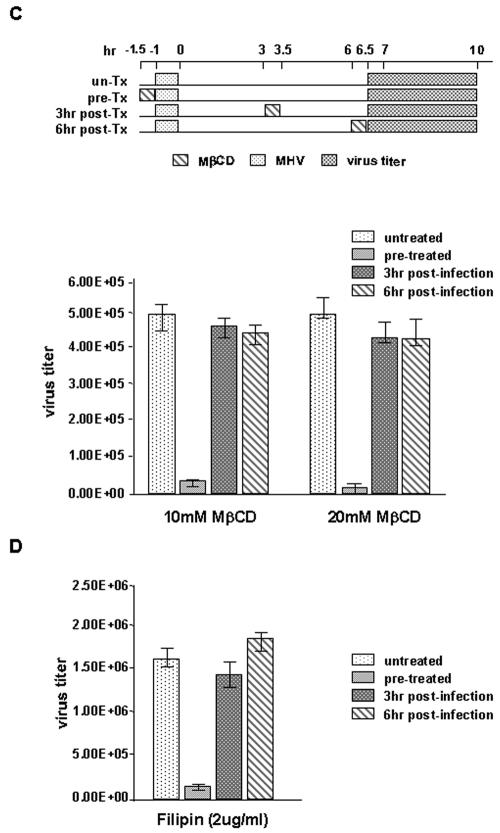
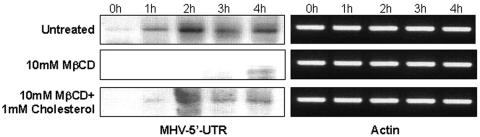
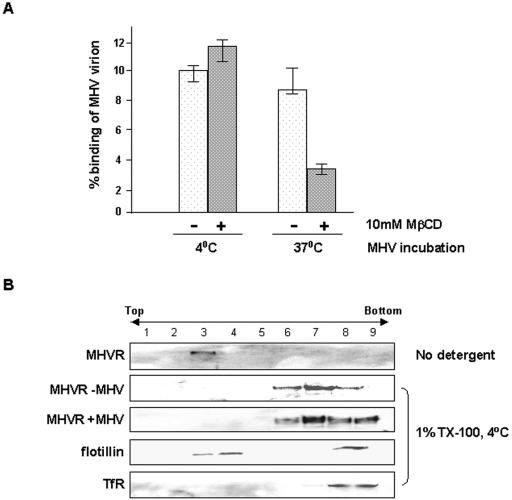
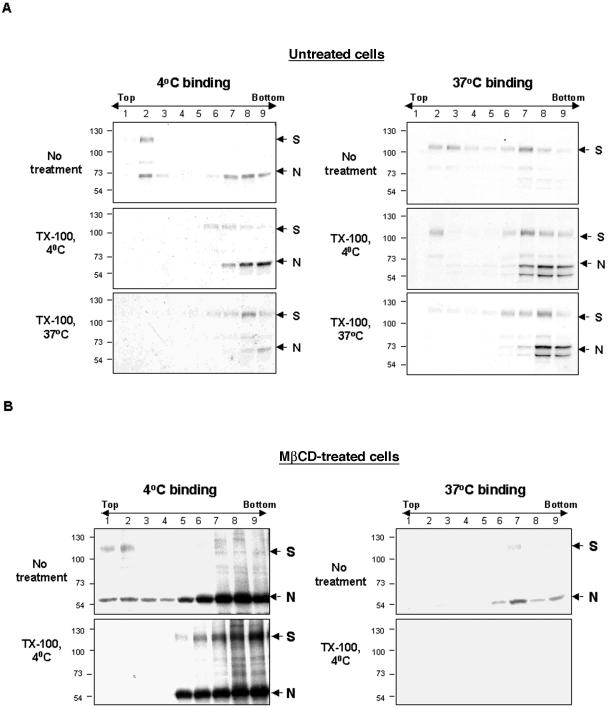
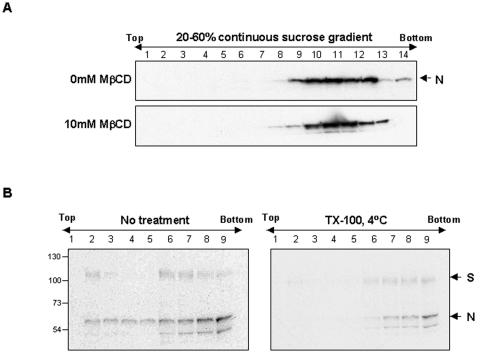
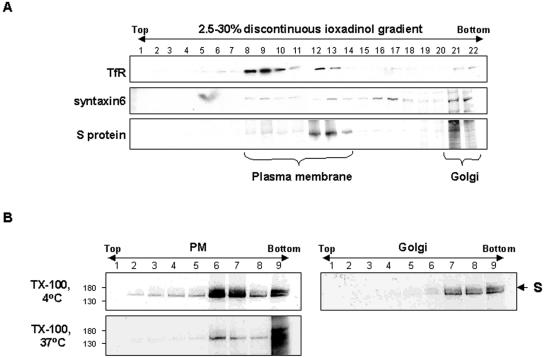
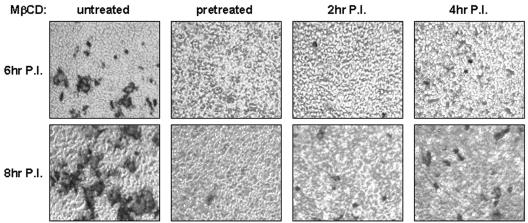
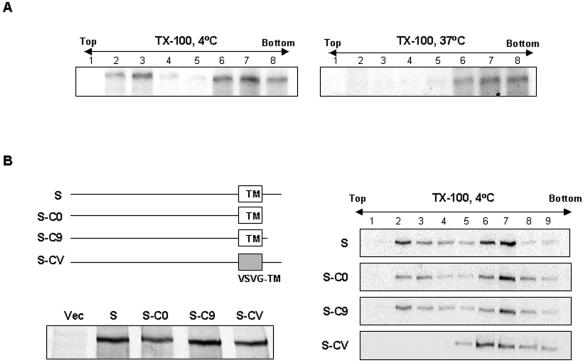
Thorp and Gallagher first reported that depletion of cholesterol inhibited virus entry and cell-cell fusion of mouse hepatitis virus (MHV), suggesting the importance of lipid rafts in MHV replication (E. B. Thorp and T. M. Gallagher, J. Virol. 78:2682-2692, 2004). However, the MHV receptor is not present in lipid rafts, and anchoring of the MHV receptor to lipid rafts did not enhance MHV infection; thus, the mechanism of lipid rafts involvement is not clear. In this study, we defined the mechanism and extent of lipid raft involvement in MHV replication. We showed that cholesterol depletion by methyl beta-cyclodextrin or filipin did not affect virus binding but reduced virus entry. Furthermore, MHV spike protein bound to nonraftraft membrane at 4 degrees C but shifted to lipid rafts at 37 degrees C, indicating a redistribution of membrane following virus binding. Thus, the lipid raft involvement in MHV entry occurs at a step following virus binding. We also found that the viral spike protein in the plasma membrane of the infected cells was associated with lipid rafts, whereas that in the Golgi membrane, where MHV matures, was not. Moreover, the buoyant density of the virion was not changed when MHV was produced from the cholesterol-depleted cells, suggesting that MHV does not incorporate lipid rafts into the virion. These results indicate that MHV release does not involve lipid rafts. However, MHV spike protein has an inherent ability to associate with lipid rafts. Correspondingly, cell-cell fusion induced by MHV was retarded by cholesterol depletion, consistent with the association of the spike protein with lipid rafts in the plasma membrane. These findings suggest that MHV entry requires specific interactions between the spike protein and lipid rafts, probably during the virus internalization step.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
