

Impaired virus control and severe CD8+ T-cell-mediated immunopathology in chimeric mice deficient in gamma interferon receptor expression on both parenchymal and hematopoietic cells
- PMID: 16014969
- PMCID: PMC1181549
- DOI: 10.1128/JVI.79.15.10073-10076.2005
Impaired virus control and severe CD8+ T-cell-mediated immunopathology in chimeric mice deficient in gamma interferon receptor expression on both parenchymal and hematopoietic cells
Abstract
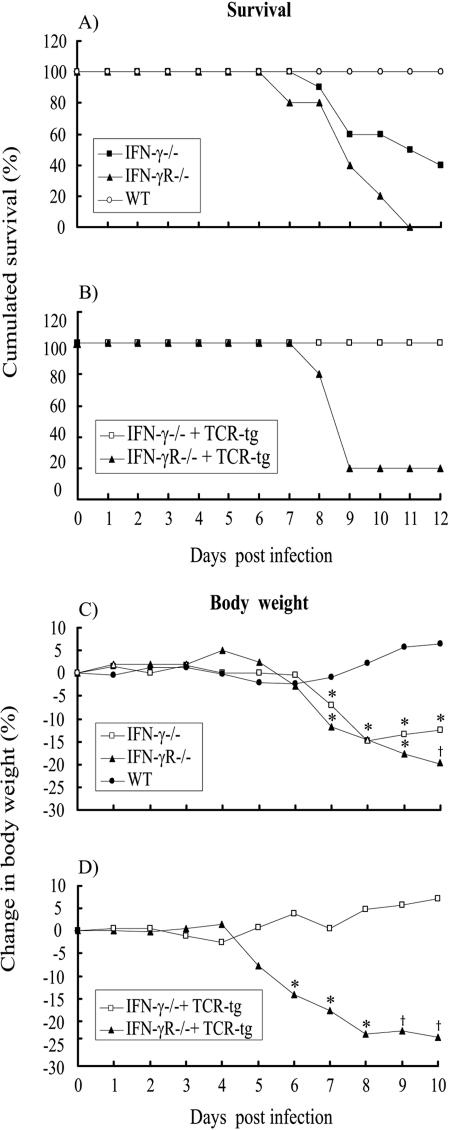
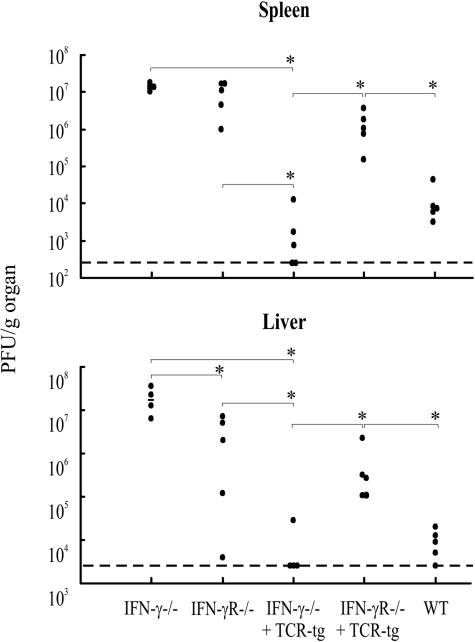
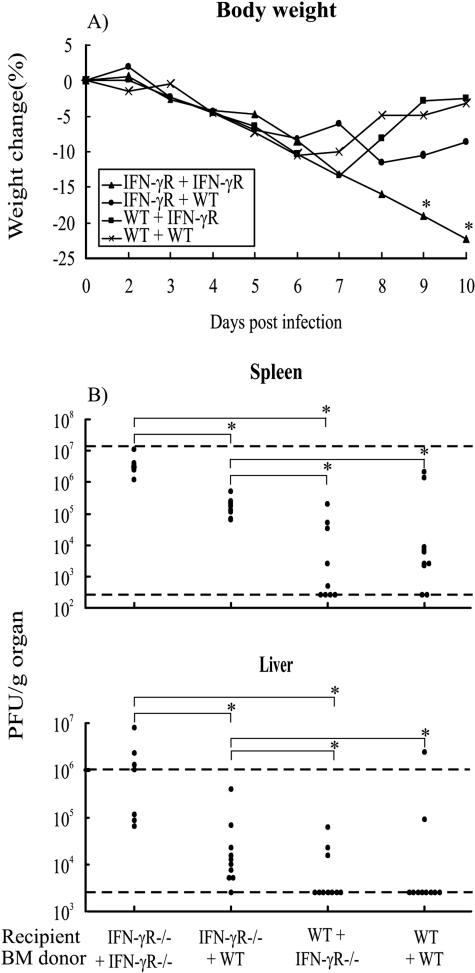
Bone marrow chimeras were used to determine the cellular target(s) for the antiviral activity of gamma interferon (IFN-gamma). By transfusing such mice with high numbers of naive virus-specific CD8(+) T cells, a system was created in which the majority of virus-specific CD8(+) T cells would be capable of responding to IFN-gamma, but expression of the relevant receptor on non-T cells could be experimentally controlled. Only when the IFN-gamma receptor is absent on both radioresistant parenchymal and bone marrow-derived cells will chimeric mice challenged with a highly invasive, noncytolytic virus completely lack the ability to control the infection and develop severe wasting disease. Further, the study shows that IFN-gamma receptor expression on parenchymal cells in the viscera is more important for virus control than IFN-gamma receptor expression on bone marrow-derived cells.
Figures
References
-
- Battegay, M., S. Cooper, A. Althage, J. Banziger, H. Hengartner, and R. M. Zinkernagel. 1991. Quantification of lymphocytic choriomeningitis virus with an immunological focus assay in 24- or 96-well plates. J. Virol. Methods 33:191-198. - PubMed
-
- Christensen, J. E., J. P. Christensen, N. N. Kristensen, N. J. Hansen, A. Stryhn, and A. R. Thomsen. 2002. Role of CD28 co-stimulation in generation and maintenance of virus-specific T cells. Int. Immunol. 14:701-711. - PubMed
-
- Christensen, J. E., D. Wodarz, J. P. Christensen, and A. R. Thomsen. 2004. Perforin and IFN-gamma do not significantly regulate the virus-specific CD8+ T cell response in the absence of antiviral effector activity. Eur. J. Immunol. 34:1389-1394. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
