

doi: 10.1101/gad.1273105.
Distinct roles for the RSC and Swi/Snf ATP-dependent chromatin remodelers in DNA double-strand break repair
Affiliations
- PMID: 16024655
- PMCID: PMC1176001
- DOI: 10.1101/gad.1273105
Item in Clipboard
Distinct roles for the RSC and Swi/Snf ATP-dependent chromatin remodelers in DNA double-strand break repair
Genes Dev.
.
Abstract
The failure of cells to repair damaged DNA can result in genomic instability and cancer. To efficiently repair chromosomal DNA lesions, the repair machinery must gain access to the damaged DNA in the context of chromatin. Here we report that both the RSC and Swi/Snf ATP-dependent chromatin-remodeling complexes play key roles in double-strand break (DSB) repair, specifically by homologous recombination (HR). RSC and Swi/Snf are each recruited to an in vivo DSB site but with distinct kinetics. We show that Swi/Snf is required earlier, at or preceding the strand invasion step of HR, while RSC is required following synapsis for completion of the recombinational repair event.
Figures
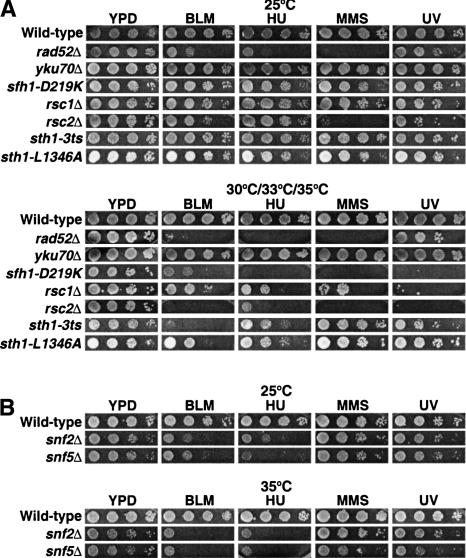
rsc and swi/snf mutants are hypersensitive to genotoxic agents. (A) Tenfold serial dilutions of wild-type (BLY76), rad52Δ (BLY772), yku70Δ (BLY773), sfh1-D219K (BLY650), rsc1Δ (BLY788), rsc2Δ (BLY662), sth1-3ts (BLY49), and sth1-L1346A (BLY491) strains were spotted onto rich YPD medium in the absence or presence of 3 mU/mL bleomycin, 50 mM hydroxyurea, 0.01% MMS, or 75 J/m2 UV light. Cells were incubated for 3–4 d at permissive (25°C) or at each of their respective semipermissive (33°C for rad52Δ, yku70Δ and sfh1-D219K; 35°C for rsc1Δ, rsc2Δ, sth1-3ts, and sth1-L1346A) temperatures; wild-type cells were grown at 30°C. (B) Tenfold serial dilutions of wild-type (BLY1), snf2Δ (BLY35), and snf5Δ (BLY3) strains were spotted onto YPD medium in the absence or presence of 3 mU/mL bleomycin, 50 mM hydroxyurea, 0.01% MMS, or 75 J/m2 UV radiation and incubated at 25°C and 35°C for 3–4 d.
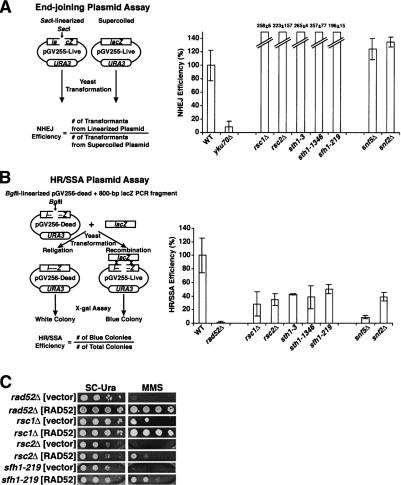
rsc and swi/snf mutants are competent for plasmid end joining but defective in repair by HR/SSA. (A) rsc and swi/snf mutants are capable of repair by plasmid end joining. Wild-type (BLY76), yku70Δ (BLY773), rsc?Δ (BLY788), rsc2Δ (BLY662), sth1-3ts (BLY49), sth1-L1346A (BLY491), sfh1-219 (BLY650), snf5Δ (BLY3), and snf2Δ (BLY16) strains transformed in parallel with the SacI-linearized or supercoiled pGV255-live plasmid were plated onto selective SC-Ura medium at 30°C for 4–5 d. Values represent the ratios of the number of transformants from linearized plasmid to the number of transformants from supercoiled plasmid, normalized to values obtained from wild-type strains. The means and standard deviations of at least three independent experiments are plotted. (B) rsc and swi/snf mutants are defective for HR/SSA. The same strains as in A and rad52Δ (BLY772) were cotransformed with the BglII-linearized pGV256-dead plasmid and an 800-bp lacZ PCR fragment amplified from pGV255-live, plated onto SC-Ura medium at 30°C for 4–5 d, and assayed for β-galactosidase activity. Values represent the ratios of blue colonies to total colonies, normalized to values obtained from wild-type strains. The means and standard deviations of at least three independent experiments are plotted. (C) RAD52 overexpression partially rescues the MMS hypersensitivity of rsc mutants. rad52Δ (BLY772), rsc1Δ (BLY788), rsc2Δ (BLY662), and sfh1-219 (BLY650) strains transformed with either pRS426:RAD52 or pRS426 alone were 10-fold serially diluted and spotted onto SC-Ura medium in the absence or presence of 0.01% MMS at 35°C (or at 33°C for sfh1-219) for 3–4 d.
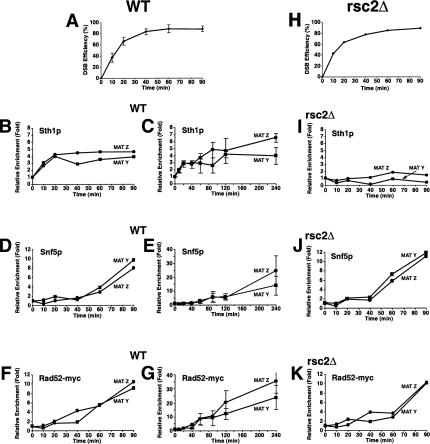
RSC and Swi/Snf are recruited to an in vivo DSB but neither is required for Rad52p's recruitment to a DSB. (A,H) Kinetics of DSB induction. Wild-type (BLY755) or rsc2Δ (BLY780) cells grown to mid-logarithmic phase in 2% raffinose at 30°C were shifted to 2% galactose to induce DSBs and genomic DNA harvested at the indicated times. The averages and standard deviations of five wild-type or three rsc2Δ mutant time-course experiments are plotted. (B,D,F) Aliquots of wild-type cells from A were processed for ChIP using anti-Sth1p, anti-Snf5p, or anti-Rad52p-13myc antibodies at the indicated times. All ChIPs were quantified by real-time PCR analysis and normalized to the time 0 values and to input DNA. Levels of CEN-proximal (MAT Y) or CEN-distal (MAT Z) DNAs immunoprecipitated by anti-Sth1p and anti-Snf5p were additionally normalized to those at the PRE1 locus. (C,E,G) Long time courses of anti-Sth1p, anti-Snf5p, or anti-Rad52-myc13p ChIP analysis show continuous recruitment of Snf5p and Rad52-myc13p to a DSB. (I–K) ChIP assays using anti-Sth1p, anti-Snf5p, or anti-Rad52-myc13p antibodies were performed on rsc2Δ (BLY780) cells as described for wild-type cells. All ChIP data are representative of at least three independent time-course experiments except for anti-Rad52-myc ChIPs, which were done twice. Error bars represent standard deviations.
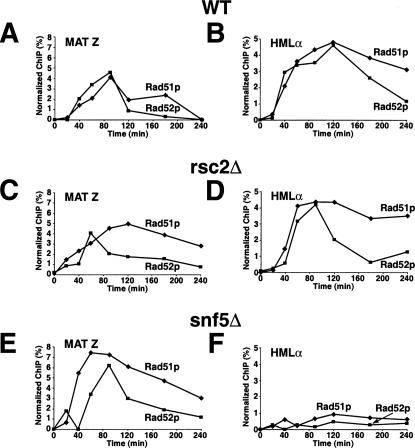
Synapsis formation between MAT and HML is defective in snf5 mutants but not rsc2 mutants. Wild-type (BLY747), rsc2Δ (BLY784), or snf5Δ (BLY786) strains carrying the HMLα donor sequence were treated with 2% galactose to induce expression of HO endonuclease, and then with 2% dextrose after 1 h to repress HO. Chromatin prepared from wild-type (A,B), rsc2Δ (C,D), or snf5Δ (E,F) cells at the indicated time points was immunoprecipitated with anti-Rad51p or anti-Rad52p antibodies. The immunoprecipitated DNAs were quantified by real-time PCR amplification using primers specific to MAT Z or HMLα regions and normalized to ACT1 promoter sequences.
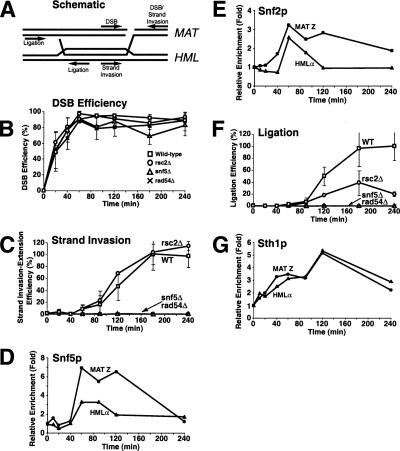
rsc and swi/snf are defective at distinct steps of HR repair. (A) Schematic diagram of an HR intermediate during mating-type switching. Primers used to monitor DSB, strand invasion-extension, and completion of repair are indicated. (B,C,F) DSBs were induced in wild-type (BLY747), rsc2Δ (BLY784), snf5Δ (BLY786), and rad54Δ (BLY804) strains as described in Figure 4. Genomic DNA was isolated at the indicated time points and monitored by quantitative real-time PCR analysis for efficiency of DSB formation (B), primer extension (C), and completion of repair (F) (see Supplemental Material for details). Primer extension and completion of repair were arbitrarily set at 100% for the highest wild-type level. All values were normalized to an ACT1 internal control. The values are the averages of three independent experiments except for experiments carried out in snf5Δ and rad54Δ strains, which were each done twice. Error bars indicate one standard deviation. (D,E,G) Chromatin was also isolated from wild-type (BLY747) strains at the indicated time points and immunoprecipitated with anti-Snf5p, anti-Snf2p, or anti-Sth1p antibodies, followed by quantitative real-time PCR analysis as described in Figure 3.
References
-
- Alexeev A., Mazin, A., and Kowalczykowski, S.C. 2003. Rad54 protein possesses chromatin-remodeling activity stimulated by the Rad51-ssDNA nucleoprotein filament. Nat. Struct. Biol. 10: 182–185. - PubMed
-
- Angus-Hill M.L., Schlichter, A., Roberts, D., Erdjument-Bromage, H., Tempst, P., and Cairns, B.R. 2001. A Rsc3/Rsc30 zinc cluster dimer reveals novel roles for the chromatin remodeler RSC in gene expression and cell cycle control. Mol. Cell 7: 741–751. - PubMed
-
- Baetz K.K., Krogan, N.J., Emili, A., Greenblatt, J., and Hieter, P. 2004. The ctf13-30/CTF13 genomic haploinsufficiency modifier screen identifies the yeast chromatin remodeling complex RSC, which is required for the establishment of sister chromatid cohesion. Mol. Cell. Biol. 24: 1232–1244. - PMC - PubMed
-
- Bennett C.B., Lewis, L.K., Karthikeyan, G., Lobachev, K.S., Jin, Y.H., Sterling, J.F., Snipe, J.R., and Resnick, M.A. 2001. Genes required for ionizing radiation resistance in yeast. Nat. Genet. 29: 426–434. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases