

Functional but abnormal adult erythropoiesis in the absence of the stem cell leukemia gene
- PMID: 16024775
- PMCID: PMC1190361
- DOI: 10.1128/MCB.25.15.6355-6362.2005
Functional but abnormal adult erythropoiesis in the absence of the stem cell leukemia gene
Abstract
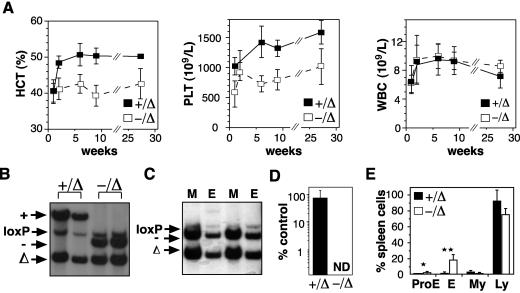
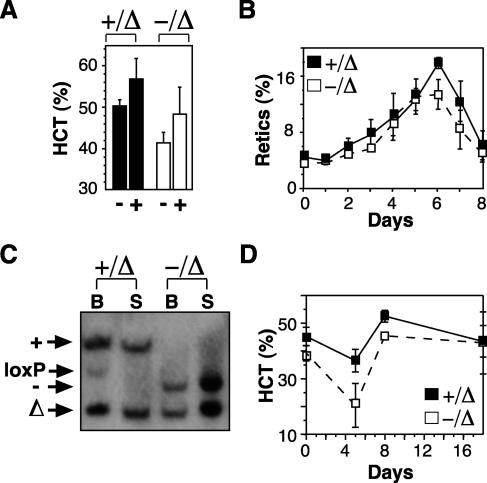
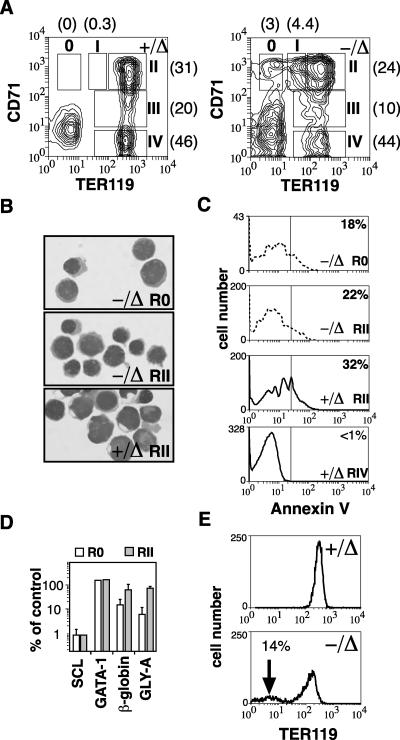
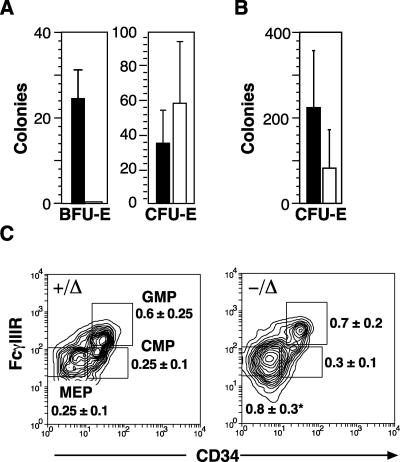
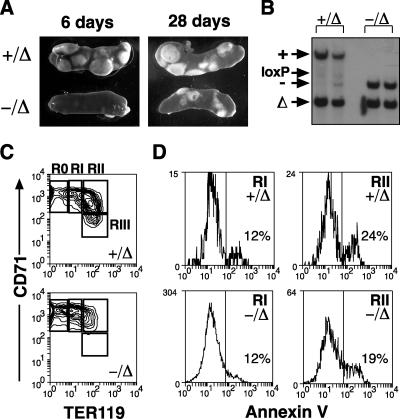
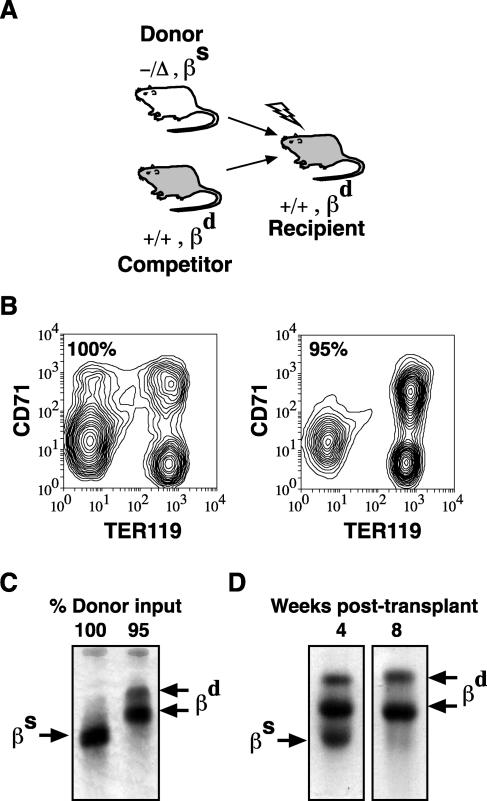
Previous studies have indicated that the stem cell leukemia gene (SCL) is essential for both embryonic and adult erythropoiesis. We have examined erythropoiesis in conditional SCL knockout mice for at least 6 months after loss of SCL function and report that SCL was important but not essential for the generation of mature red blood cells. Although SCL-deleted mice were mildly anemic with increased splenic erythropoiesis, they responded appropriately to endogenous erythropoietin and hemolytic stress, a measure of late erythroid progenitors. However, SCL was more important for the proliferation of early erythroid progenitors because the predominant defects in SCL-deleted erythropoiesis were loss of in vitro growth of the burst-forming erythroid unit and an in vivo growth defect revealed by transplant assays. With respect to erythroid maturation, SCL-deleted proerythroblasts could generate more mature erythroblasts and circulating red blood cells. However, SCL was required for normal expression of TER119, one of the few proposed target genes of SCL. The unexpected finding that SCL-independent erythropoiesis can proceed in the adult suggests that alternate factors can replace the essential functions of SCL and raises the possibility that similar mechanisms also explain the relatively minor defects previously observed in SCL-null hematopoietic stem cells.
Figures
Similar articles
-
Control of erythroid cell production via caspase-mediated cleavage of transcription factor SCL/Tal-1.Cell Death Differ. 2003 Aug;10(8):905-13. doi: 10.1038/sj.cdd.4401255. Cell Death Differ. 2003. PMID: 12867998
-
Impaired in vitro erythropoiesis following deletion of the Scl (Tal1) +40 enhancer is largely compensated for in vivo despite a significant reduction in expression.Mol Cell Biol. 2013 Mar;33(6):1254-66. doi: 10.1128/MCB.01525-12. Epub 2013 Jan 14. Mol Cell Biol. 2013. PMID: 23319051 Free PMC article.
-
Haematopoietic stem cells retain long-term repopulating activity and multipotency in the absence of stem-cell leukaemia SCL/tal-1 gene.Nature. 2003 Jan 30;421(6922):547-51. doi: 10.1038/nature01345. Epub 2003 Jan 19. Nature. 2003. PMID: 12540851
-
Concise review: Blood relatives: formation and regulation of hematopoietic stem cells by the basic helix-loop-helix transcription factors stem cell leukemia and lymphoblastic leukemia-derived sequence 1.Stem Cells. 2012 Jun;30(6):1053-8. doi: 10.1002/stem.1093. Stem Cells. 2012. PMID: 22593015 Review.
-
The SCL/TAL1 gene: roles in normal and malignant haematopoiesis.Bioessays. 1997 Jul;19(7):607-13. doi: 10.1002/bies.950190711. Bioessays. 1997. PMID: 9230693 Review.
Cited by
-
Constitutive phosphorylation of GATA-1 at serine²⁶ attenuates the colony-forming activity of erythrocyte-committed progenitors.PLoS One. 2013 May 22;8(5):e64269. doi: 10.1371/journal.pone.0064269. Print 2013. PLoS One. 2013. PMID: 23717580 Free PMC article.
-
T-cell acute leukemia 1 (TAL1) regulation of erythropoietin receptor and association with excessive erythrocytosis.J Biol Chem. 2012 Oct 26;287(44):36720-31. doi: 10.1074/jbc.M112.378398. Epub 2012 Sep 16. J Biol Chem. 2012. PMID: 22982397 Free PMC article.
-
Genome-wide identification of TAL1's functional targets: insights into its mechanisms of action in primary erythroid cells.Genome Res. 2010 Aug;20(8):1064-83. doi: 10.1101/gr.104935.110. Epub 2010 Jun 21. Genome Res. 2010. PMID: 20566737 Free PMC article.
-
Stress hematopoiesis is regulated by the Krüppel-like transcription factor ZBP-89.Stem Cells. 2014 Mar;32(3):791-801. doi: 10.1002/stem.1598. Stem Cells. 2014. PMID: 24549639 Free PMC article.
-
Hematopoietic transcriptional mechanisms: from locus-specific to genome-wide vantage points.Exp Hematol. 2014 Aug;42(8):618-29. doi: 10.1016/j.exphem.2014.05.004. Epub 2014 May 9. Exp Hematol. 2014. PMID: 24816274 Free PMC article. Review.
References
-
- Anderson, K. P., S. C. Crable, and J. B. Lingrel. 2000. The GATA-E box-GATA motif in the EKLF promoter is required for in vivo expression. Blood 95:1652-1655. - PubMed
-
- Auffray, I., S. Marfatia, K. de Jong, G. Lee, C. H. Huang, C. Paszty, M. J. Tanner, N. Mohandas, and J. A. Chasis. 2001. Glycophorin A dimerization and band 3 interaction during erythroid membrane biogenesis: in vivo studies in human glycophorin A transgenic mice. Blood 97:2872-2878. - PubMed
-
- Begley, C. G., P. D. Aplan, M. P. Davey, K. Nakahara, K. Tchorz, J. Kurtzberg, M. S. Hershfield, B. F. Haynes, D. I. Cohen, T. A. Waldmann, and I. R. Kirsch. 1989. Chromosomal translocation in a human leukemic stem-cell line disrupts the T-cell antigen receptor delta-chain diversity region and results in a previously unreported fusion transcript. Proc. Natl. Acad. Sci. USA 86:2031-2035. - PMC - PubMed
-
- Condorelli, G., L. Vitelli, M. Valtieri, I. Marta, E. Montesoro, V. Lulli, R. Baer, and C. Peschle. 1995. Coordinate expression and developmental role of Id2 protein and TAL1/E2A heterodimer in erythroid progenitor differentiation. Blood 86:164-175. - PubMed
-
- Curtis, D. J., M. A. Hall, L. J. Van Stekelenburg, L. Robb, S. M. Jane, and C. G. Begley. 2004. SCL is required for normal function of short-term repopulating hematopoietic stem cells. Blood 103:3342-3348. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous