

Compound genetic ablation of nidogen 1 and 2 causes basement membrane defects and perinatal lethality in mice
- PMID: 16024816
- PMCID: PMC1190363
- DOI: 10.1128/MCB.25.15.6846-6856.2005
Compound genetic ablation of nidogen 1 and 2 causes basement membrane defects and perinatal lethality in mice
Abstract
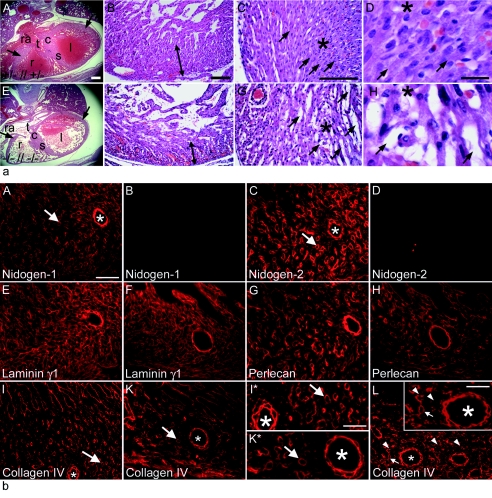
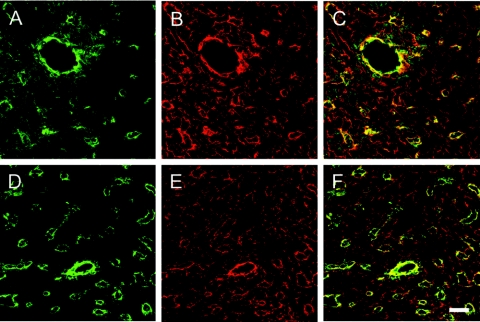
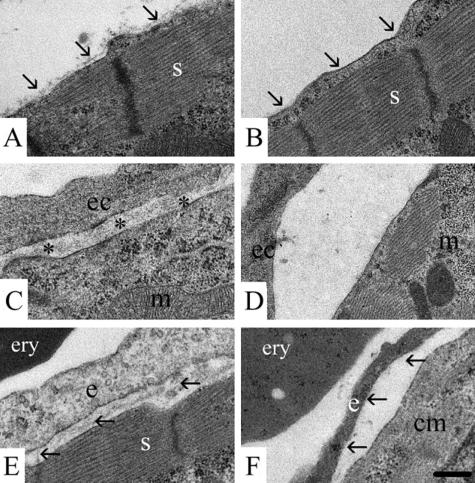
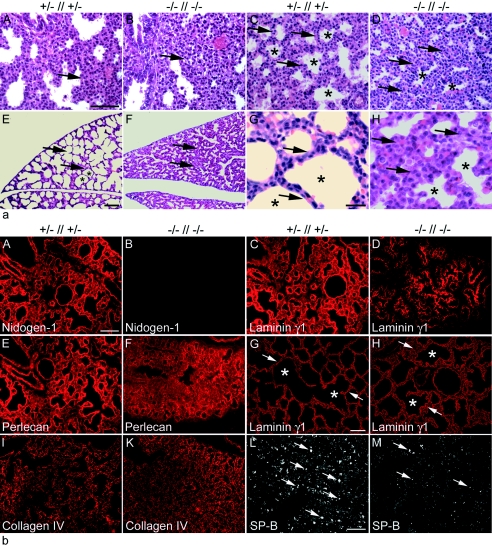
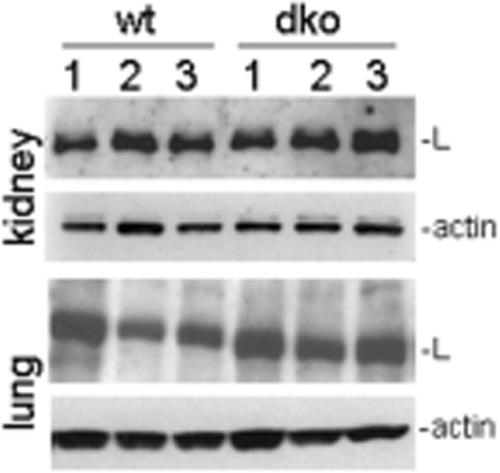
Nidogen 1 and 2 are basement membrane glycoproteins, and previous biochemical and functional studies indicate that they may play a crucial role in basement membrane assembly. While they show a divergent expression pattern in certain adult tissues, both have a similar distribution during development. Gene knockout studies in mice demonstrated that the loss of either isoform has no effect on basement membrane formation and organ development, suggesting complementary functions. Here, we show that this is indeed the case. Deficiency of both nidogens in mice resulted in perinatal lethality. Nidogen 1 and 2 do not appear to be crucial in establishing tissue architecture during organ development; instead, they are essential for late stages of lung development and for maintenance and/or integrity of cardiac tissue. These organ defects are not compatible with postnatal survival. Ultrastructural analysis suggests that the phenotypes directly result from basement membrane changes. However, despite the ubiquitous presence of nidogens in basement membranes, defects do not occur in all tissues or in all basement membranes, suggesting a varying spectrum of roles for nidogens in the basement membrane.
Figures


References
-
- Arikawa-Hirasawa, E., H. Watanabe, H. Takami, J. R. Hassell, and Y. Yamada. 1999. Perlecan is essential for cartilage and cephalic development. Nat. Genet. 23:354-358. - PubMed
-
- Aumailley, M., C. Battaglia, U. Mayer, R. Nischt, R. Timpl, and J. W. Fox. 1993. Nidogen mediates the formation of ternary complexes of basement membrane components. Kidney Int. 43:7-12. - PubMed
-
- Bader, B. L., H. Rayburn, D. Crowley, and R. Hynes. 1998. Extensive vasculogenesis, angiogenesis, and organogenesis precede lethality in mice lacking αv integrins. Cell 95:507-519. - PubMed
-
- Breitkreutz, D., N. Mirancea, C. Schmidt, R. Beck, U. Werner, H.-J. Stark, M. Gerl, and N. Fusenig. 2004. Inhibition of basement membrane formation by a nidogen-binding laminin γ1-chain fragment in human skin-organotypic cocultures. J. Cell Sci. 117:2611-2622. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases