

Inhibition of Henipavirus fusion and infection by heptad-derived peptides of the Nipah virus fusion glycoprotein
- PMID: 16026621
- PMCID: PMC1208959
- DOI: 10.1186/1743-422X-2-57
Inhibition of Henipavirus fusion and infection by heptad-derived peptides of the Nipah virus fusion glycoprotein
Abstract
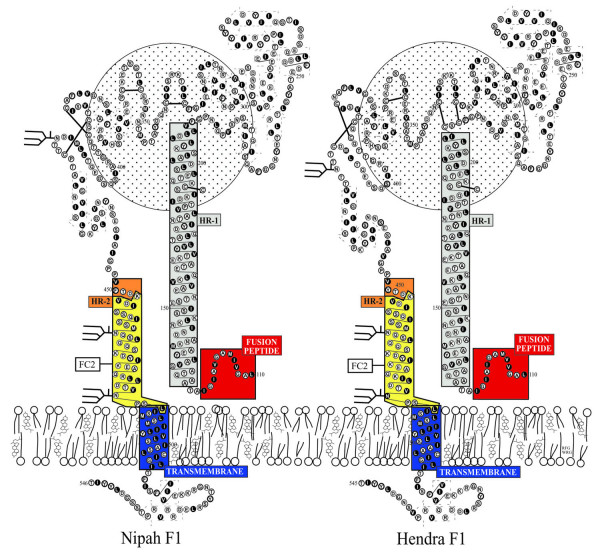
Background: The recent emergence of four new members of the paramyxovirus family has heightened the awareness of and re-energized research on new and emerging diseases. In particular, the high mortality and person to person transmission associated with the most recent Nipah virus outbreaks, as well as the very recent re-emergence of Hendra virus, has confirmed the importance of developing effective therapeutic interventions. We have previously shown that peptides corresponding to the C-terminal heptad repeat (HR-2) of the fusion envelope glycoprotein of Hendra virus and Nipah virus were potent inhibitors of both Hendra virus and Nipah virus-mediated membrane fusion using recombinant expression systems. In the current study, we have developed shorter, second generation HR-2 peptides which include a capped peptide via amidation and acetylation and two poly(ethylene glycol)-linked (PEGylated) peptides, one with the PEG moity at the C-terminus and the other at the N-terminus. Here, we have evaluated these peptides as well as the corresponding scrambled peptide controls in Nipah virus and Hendra virus-mediated membrane fusion and against infection by live virus in vitro.
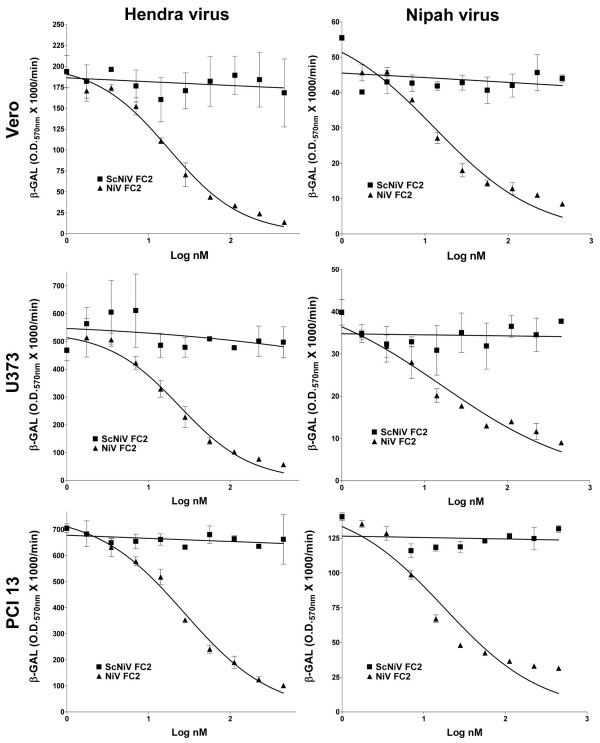
Results: Unlike their predecessors, the second generation HR-2 peptides exhibited high solubility and improved synthesis yields. Importantly, both Nipah virus and Hendra virus-mediated fusion as well as live virus infection were potently inhibited by both capped and PEGylated peptides with IC50 concentrations similar to the original HR-2 peptides, whereas the scrambled modified peptides had no inhibitory effect. These data also indicate that these chemical modifications did not alter the functional properties of the peptides as inhibitors.
Conclusion: Nipah virus and Hendra virus infection in vitro can be potently blocked by specific HR-2 peptides. The improved synthesis and solubility characteristics of the second generation HR-2 peptides will facilitate peptide synthesis for pre-clinical trial application in an animal model of Henipavirus infection. The applied chemical modifications are also predicted to increase the serum half-life in vivo and should increase the chance of success in the development of an effective antiviral therapy.
Figures
References
-
- Murray K, Selleck P, Hooper P, Hyatt A, Gould A, Gleeson L, Westbury H, Hiley L, Selvey L, Rodwell B, et al. A morbillivirus that caused fatal disease in horses and humans. Science. 1995;268:94–97. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
