

CD4-CD8- T cells control intracellular bacterial infections both in vitro and in vivo
- PMID: 16027239
- PMCID: PMC2212999
- DOI: 10.1084/jem.20050569
CD4-CD8- T cells control intracellular bacterial infections both in vitro and in vivo
Abstract
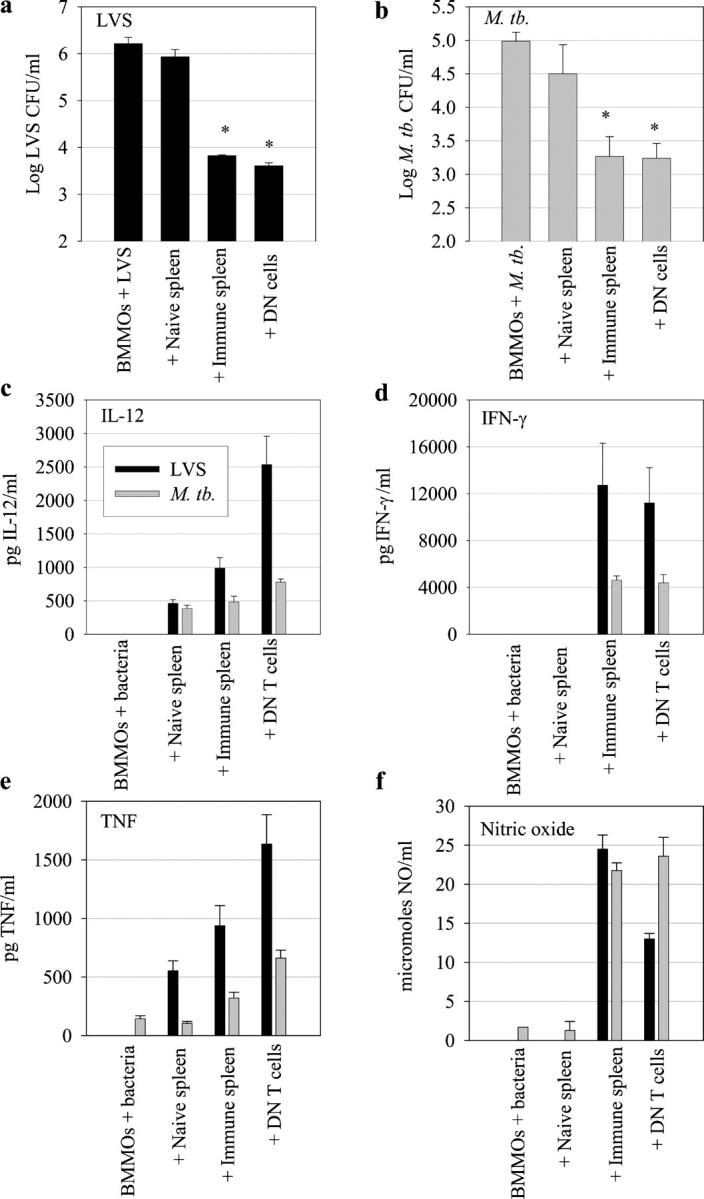
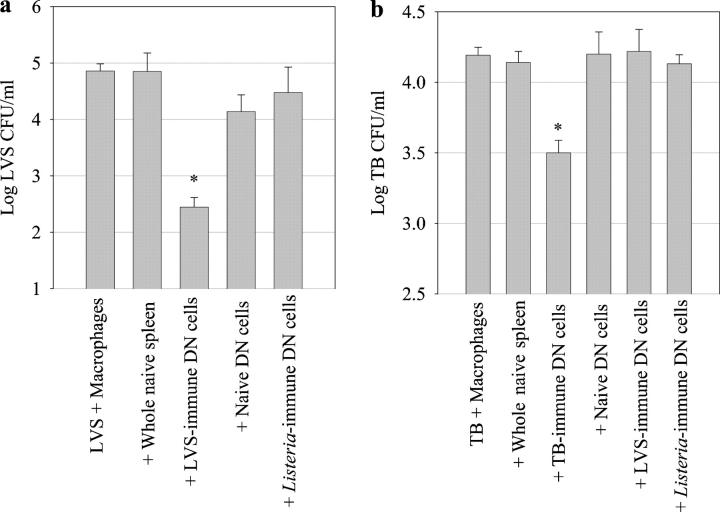
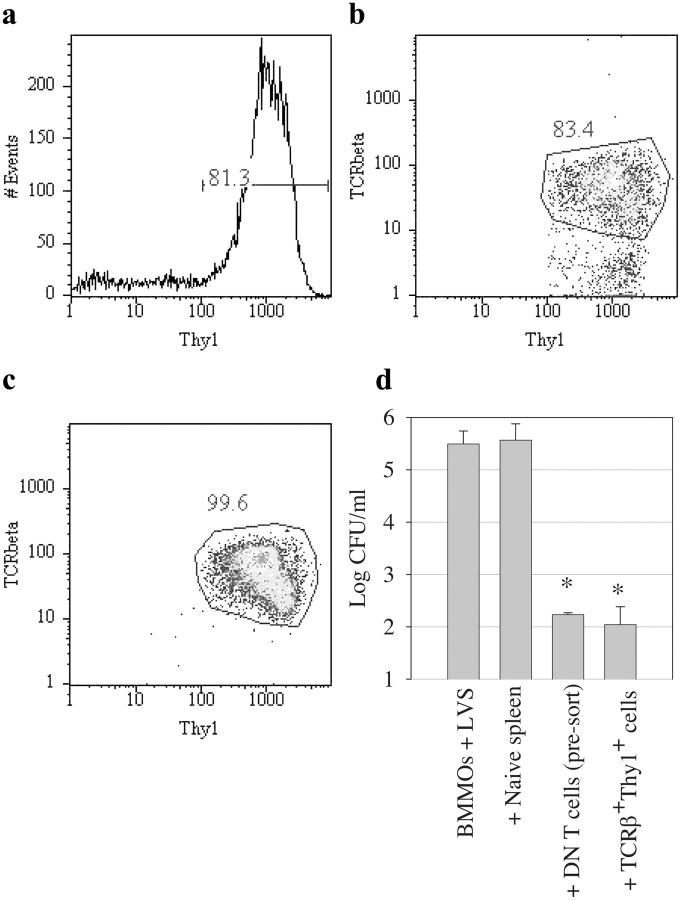
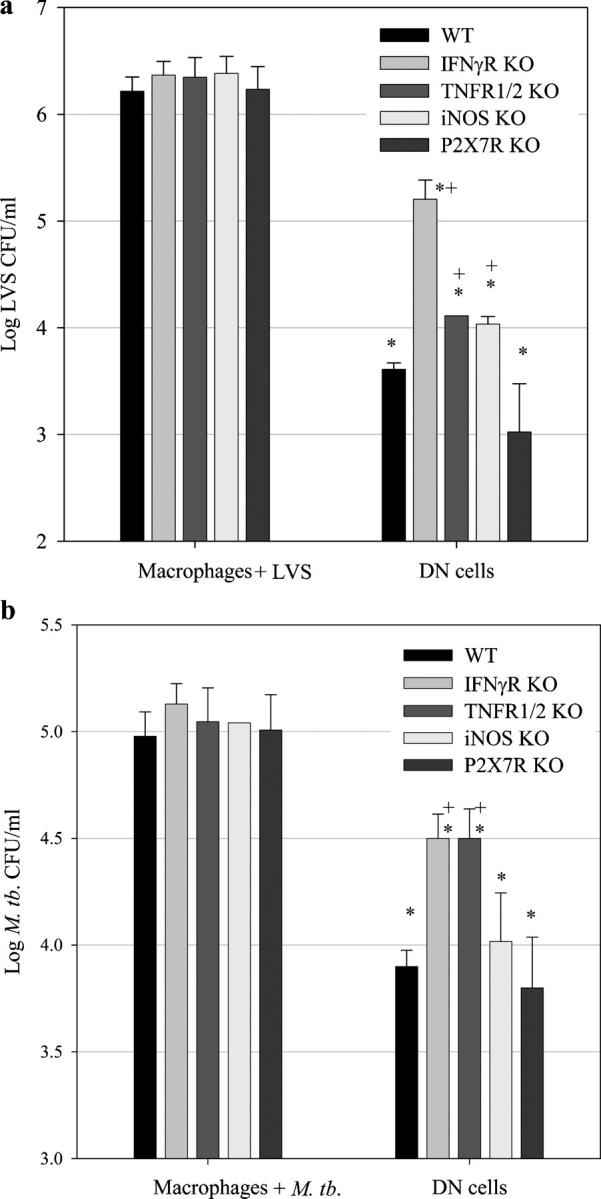
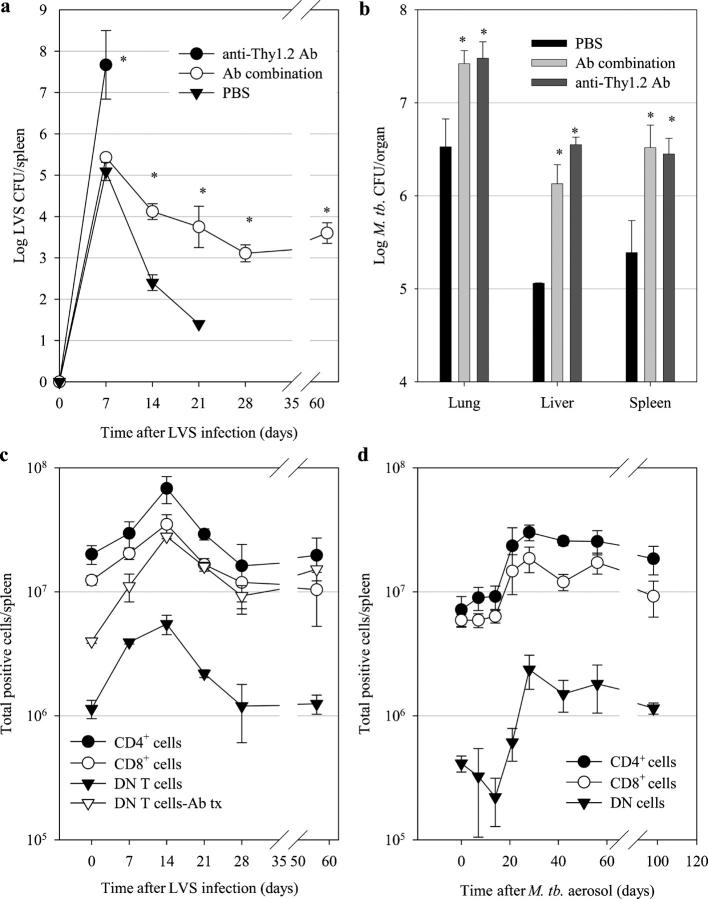
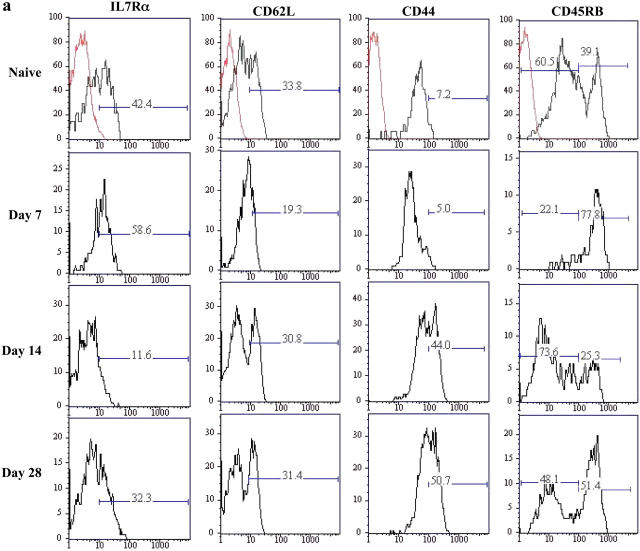
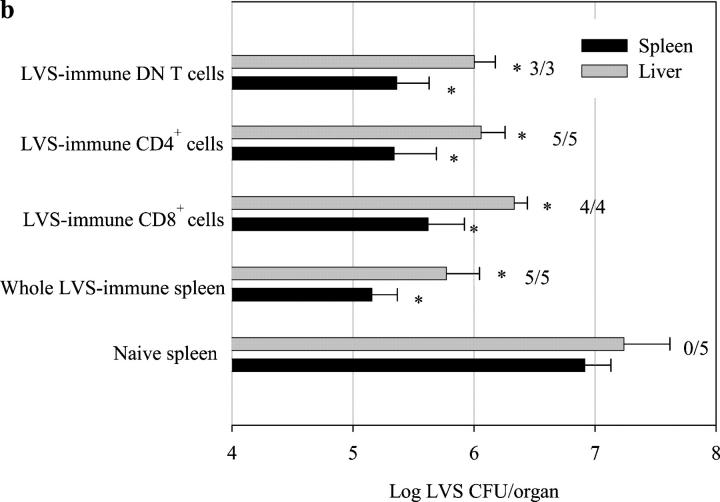
Memory T cells, including the well-known CD4(+) and CD8(+) T cells, are central components of the acquired immune system and are the basis for successful vaccination. After infection, CD4(+) and CD8(+) T cells expand into effector cells, and then differentiate into long-lived memory cells. We show that a rare population of CD4(-)CD8(-)CD3(+)alphabeta(+)gammadelta(-)NK1.1(-) T cells has similar functions. These cells potently and specifically inhibit the growth of the intracellular bacteria Mycobacterium tuberculosis (M. tb.) or Francisella tularensis Live Vaccine Strain (LVS) in macrophages in vitro, promote survival of mice infected with these organisms in vivo, and adoptively transfer immunity to F. tularensis LVS. Furthermore, these cells expand in the spleens of mice infected with M. tb. or F. tularensis LVS, and then acquire a memory cell phenotype. Thus, CD4(-)CD8(-) T cells have a role in the control of intracellular infection and may contribute to successful vaccination.
Figures
Similar articles
-
Loss of either CD4+ or CD8+ T cells does not affect the magnitude of protective immunity to an intracellular pathogen, Francisella tularensis strain LVS.J Immunol. 1996 Dec 1;157(11):5042-8. J Immunol. 1996. PMID: 8943413
-
Involvement of NK1+ CD4- CD8- alphabeta T cells and endogenous IL-4 in non-MHC-restricted rejection of embryonal carcinoma in genetically resistant mice.J Immunol. 1997 Jun 1;158(11):5338-48. J Immunol. 1997. PMID: 9164954
-
Susceptibility to secondary Francisella tularensis live vaccine strain infection in B-cell-deficient mice is associated with neutrophilia but not with defects in specific T-cell-mediated immunity.Infect Immun. 2001 Jan;69(1):194-203. doi: 10.1128/IAI.69.1.194-203.2001. Infect Immun. 2001. PMID: 11119506 Free PMC article.
-
[Novel vaccines against M. tuberculosis].Kekkaku. 2006 Dec;81(12):745-51. Kekkaku. 2006. PMID: 17240920 Review. Japanese.
-
Porcine T lymphocytes and NK cells--an update.Dev Comp Immunol. 2009 Mar;33(3):310-20. doi: 10.1016/j.dci.2008.06.003. Epub 2008 Jul 2. Dev Comp Immunol. 2009. PMID: 18601948 Review.
Cited by
-
Development of functional and molecular correlates of vaccine-induced protection for a model intracellular pathogen, F. tularensis LVS.PLoS Pathog. 2012 Jan;8(1):e1002494. doi: 10.1371/journal.ppat.1002494. Epub 2012 Jan 19. PLoS Pathog. 2012. PMID: 22275868 Free PMC article.
-
A defined O-antigen polysaccharide mutant of Francisella tularensis live vaccine strain has attenuated virulence while retaining its protective capacity.Infect Immun. 2007 May;75(5):2591-602. doi: 10.1128/IAI.01789-06. Epub 2007 Feb 12. Infect Immun. 2007. PMID: 17296751 Free PMC article.
-
Identification of a dominant CD4 T cell epitope in the membrane lipoprotein Tul4 from Francisella tularensis LVS.Mol Immunol. 2009 May;46(8-9):1830-8. doi: 10.1016/j.molimm.2009.01.008. Epub 2009 Feb 23. Mol Immunol. 2009. PMID: 19233475 Free PMC article.
-
Interleukin-6 is essential for primary resistance to Francisella tularensis live vaccine strain infection.Infect Immun. 2013 Feb;81(2):585-97. doi: 10.1128/IAI.01249-12. Epub 2012 Dec 10. Infect Immun. 2013. PMID: 23230288 Free PMC article.
-
Immune lymphocytes halt replication of Francisella tularensis LVS within the cytoplasm of infected macrophages.Sci Rep. 2020 Jul 21;10(1):12023. doi: 10.1038/s41598-020-68798-2. Sci Rep. 2020. PMID: 32694562 Free PMC article.
References
-
- Flynn, J.L., and J. Chan. 2001. Immunology of tuberculosis. Annu. Rev. Immunol. 19:93–129. - PubMed
-
- Dennis, D.T., T.V. Inglesby, D.A. Henderson, J.G. Bartlett, M.S. Ascher, E. Eitzen, A.D. Fine, A.M. Friedlander, J. Hauer, M. Layton, et al. 2001. Tularemia as a biological weapon: medical and public health management. JAMA. 285:2763–2773. - PubMed
-
- Schaible, U.E., H.L. Collins, and S.H. Kaufmann. 1999. Confrontation between intracellular bacteria and the immune system. Adv. Immunol. 71:267–377. - PubMed
-
- Elkins, K.L., S.C. Cowley, and C.M. Bosio. 2003. Innate and adaptive immune responses to an intracellular bacterium, Francisella tularensis live vaccine strain. Microbes Infect. 5:135–142. - PubMed
-
- Yee, D., T.R. Rhinehart-Jones, and K.L. Elkins. 1996. Loss of either CD4+ or CD8+ T cells does not affect the magnitude of protective immunity to an intracellular pathogen, Francisella tularensis strain LVS. J. Immunol. 157:5042–5048. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
