

Stimulation of mammalian translation initiation factor eIF4A activity by a small molecule inhibitor of eukaryotic translation
- PMID: 16030146
- PMCID: PMC1176247
- DOI: 10.1073/pnas.0504249102
Stimulation of mammalian translation initiation factor eIF4A activity by a small molecule inhibitor of eukaryotic translation
Abstract
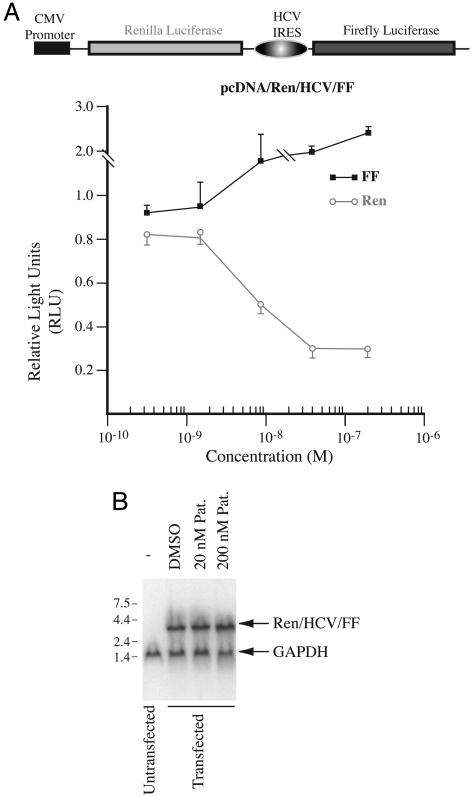
RNA helicases are the largest group of enzymes in eukaryotic RNA metabolism. The DEXD/H-box putative RNA helicases form the helicase superfamily II, whose members are defined by seven highly conserved amino acid motifs, making specific targeting of selected members a challenging pharmacological problem. The translation initiation factor eIF4A is the prototypical DEAD-box RNA helicase that works in conjunction with eIF4B and eIF4H and as a subunit of eIF4F to prepare the mRNA template for ribosome binding, possibly by unwinding the secondary structure proximal to the 5' m7GpppN cap structure. We report the identification and characterization of a small molecule inhibitor of eukaryotic translation initiation that acts in an unusual manner by stimulating eIF4A-associated activities. Our results suggest that proper control of eIF4A helicase activity is necessary for efficient ribosome binding and demonstrate the feasibility of selectively targeting DEAD-box RNA helicases with small molecules.
Figures
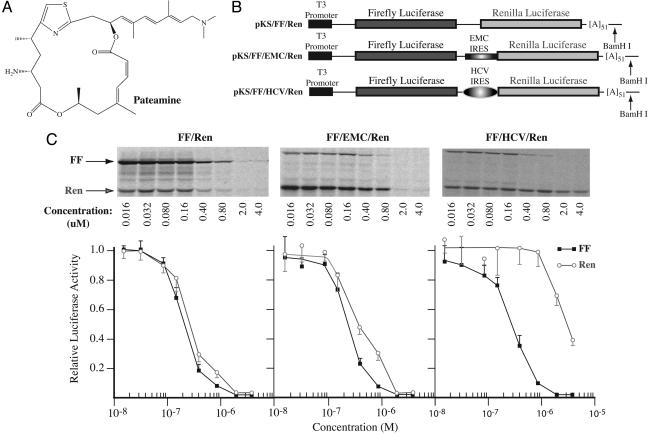
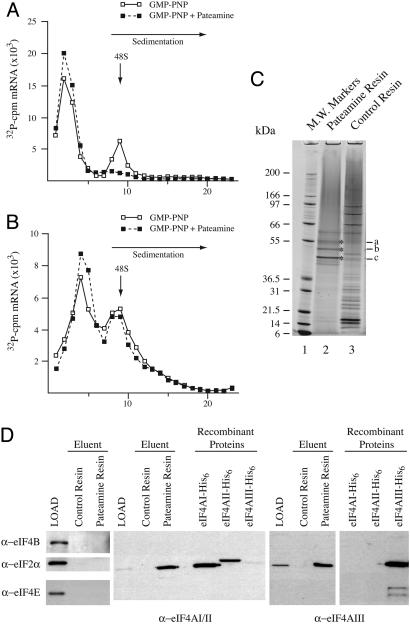
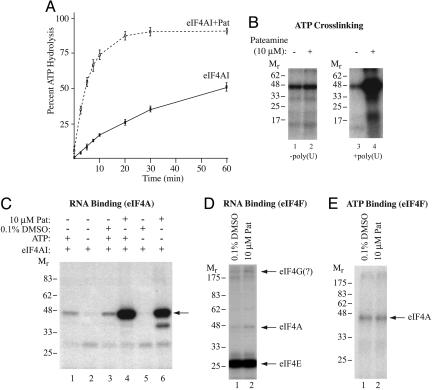
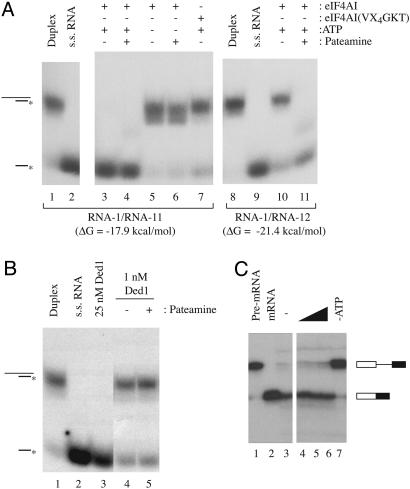
References
-
- Hershey, J. W. B. & Merrick, W. C. (2000) Initiation of Protein Synthesis (Cold Spring Habor Lab. Press, Plainview, NY).
-
- Wilson, J. E., Pestova, T. V., Hellen, C. U. & Sarnow, P. (2000) Cell 102, 511-520. - PubMed
-
- Grifo, J. A., Abramson, R. D., Satler, C. A. & Merrick, W. C. (1984) J. Biol. Chem. 259, 8648-8654. - PubMed
-
- Rogers, G. W., Jr., Richter, N. J. & Merrick, W. C. (1999) J. Biol. Chem. 274, 12236-12244. - PubMed
-
- Richter-Cook, N. J., Dever, T. E., Hensold, J. O. & Merrick, W. C. (1998) J. Biol. Chem. 273, 7579-7587. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
