

Amino acids in positions 48, 52, and 73 differentiate the substrate specificities of the highly homologous chlorocatechol 1,2-dioxygenases CbnA and TcbC
- PMID: 16030237
- PMCID: PMC1196051
- DOI: 10.1128/JB.187.15.5427-5436.2005
Amino acids in positions 48, 52, and 73 differentiate the substrate specificities of the highly homologous chlorocatechol 1,2-dioxygenases CbnA and TcbC
Erratum in
- J Bacteriol. 2005 Sep;187(18):6602
Abstract
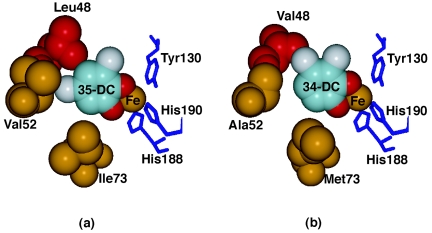
Chlorocatechol 1,2-dioxygenase (CCD) is the first-step enzyme of the chlorocatechol ortho-cleavage pathway, which plays a central role in the degradation of various chloroaromatic compounds. Two CCDs, CbnA from the 3-chlorobenzoate-degrader Ralstonia eutropha NH9 and TcbC from the 1,2,4-trichlorobenzene-degrader Pseudomonas sp. strain P51, are highly homologous, having only 12 different amino acid residues out of identical lengths of 251 amino acids. But CbnA and TcbC are different in substrate specificities against dichlorocatechols, favoring 3,5-dichlorocatechol (3,5-DC) and 3,4-dichlorocatechol (3,4-DC), respectively. A study of chimeric mutants constructed from the two CCDs indicated that the N-terminal parts of the enzymes were responsible for the difference in the substrate specificities. Site-directed mutagenesis studies further identified the amino acid in position 48 (Leu in CbnA and Val in TcbC) as critical in differentiating the substrate specificities of the enzymes, which agreed well with molecular modeling of the two enzymes. Mutagenesis studies also demonstrated that Ile-73 of CbnA and Ala-52 of TcbC were important for their high levels of activity towards 3,5-DC and 3,4-DC, respectively. The importance of Ile-73 for 3,5-DC specificity determination was also shown with other CCDs such as TfdC from Burkholderia sp. NK8 and TfdC from Alcaligenes sp. CSV90 (identical to TfdC from R. eutropha JMP134), which convert 3,5-DC preferentially. Together with amino acid sequence comparisons indicating high conservation of Leu-48 and Ile-73 among CCDs, these results suggested that TcbC of strain P51 had diverged from other CCDs to be adapted to conversion of 3,4-DC.
Figures

References
-
- Bhat, M. A., T. Ishida, K. Horiike, C. S. Vaidyanathan, and M. Nozaki. 1993. Purification of 3,5-dichlorocatechol 1,2-dioxygenase, a nonheme iron dioxygenase and a key enzyme in the biodegradation of a herbicide, 2,4-dichlorophenoxyacetic acid (2,4-D), from Pseudomonas cepacia CSV90. Arch. Biochem. Biophys. 300:738-746. - PubMed
-
- Bollag, J.-M., C. S. Helling, and M. Alexander. 1968. 2,4-D metabolism. Enzymatic hydroxylation of chlorinated phenols. J. Agr. Food Chem. 16:826-828.
-
- Broderick, J. B., and T. V. O'Halloran. 1991. Overproduction, purification, and characterization of chlorocatechol dioxygenase, a non-heme iron dioxygenase with broad substrate tolerance. Biochemistry 30:7349-7358. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
