

Expression of sucrose synthase genes involved in enhanced elongation of pondweed (Potamogeton distinctus) turions under anoxia
- PMID: 16033779
- PMCID: PMC4247035
- DOI: 10.1093/aob/mci220
Expression of sucrose synthase genes involved in enhanced elongation of pondweed (Potamogeton distinctus) turions under anoxia
Abstract
Background and aims: Overwintering buds (turions) of the monocot aquatic pondweed species (Potamogeton distinctus) are highly tolerant to anoxic stress. Sucrose metabolism accompanied by enhanced activity of sucrose synthase (SuSy) operates actively during anaerobic elongation of pondweed turions. The aim of this study is to isolate SuSy genes from the turions and to investigate their transcriptional changes in response to anoxia and other stimuli.
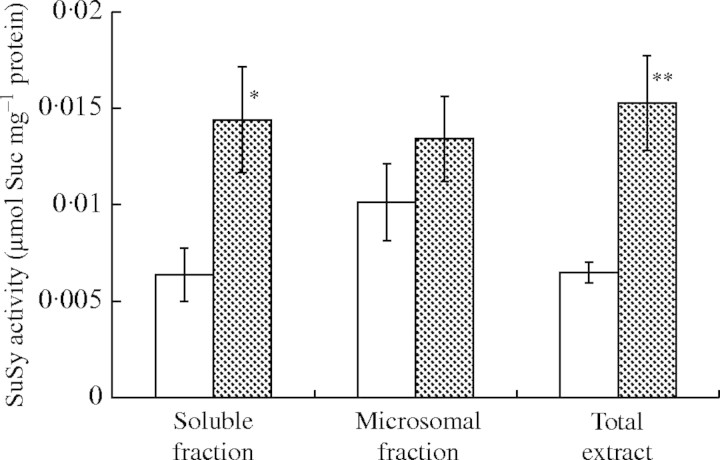
Methods: SuSy genes were isolated from pondweed turions by PCR methods and transcript levels of SuSy genes were examined in response to anoxia, sugars and plant hormones. In addition, the effects of anoxia on SuSy activity were examined both in the soluble fraction and in the microsomal fraction.
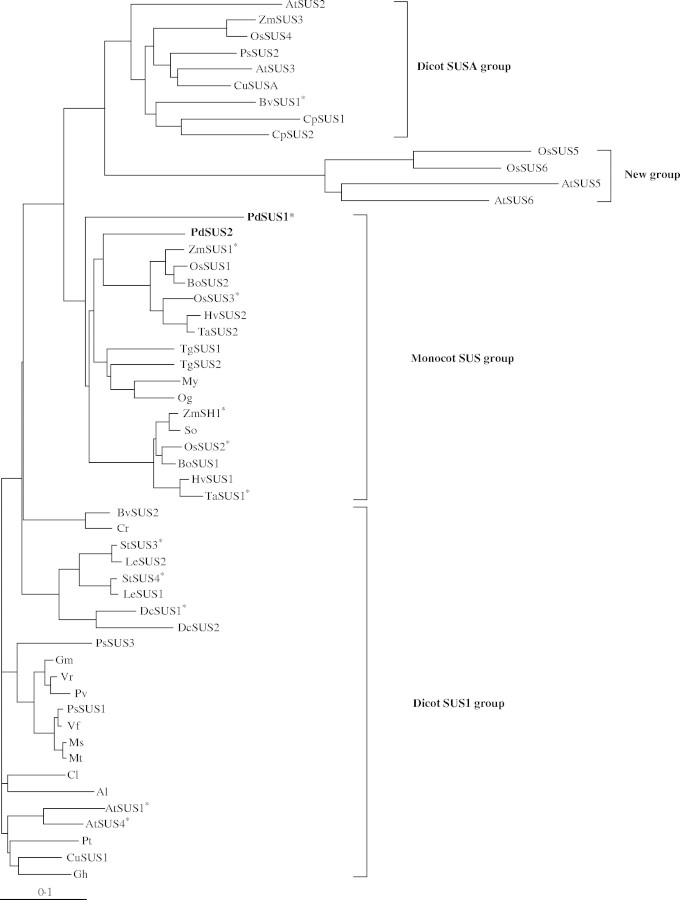
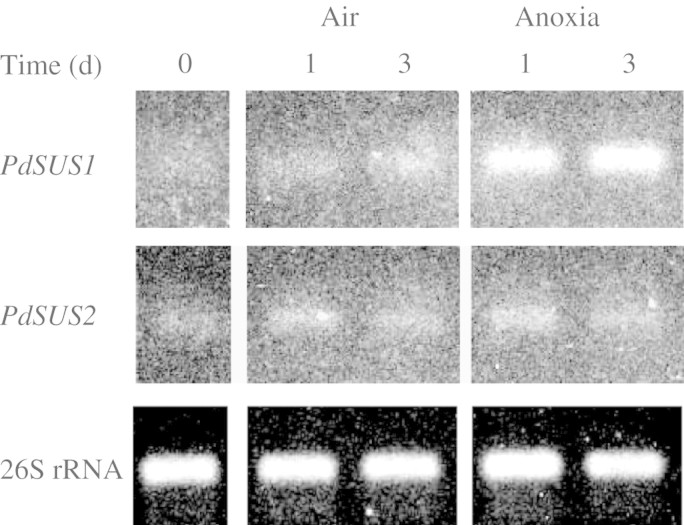
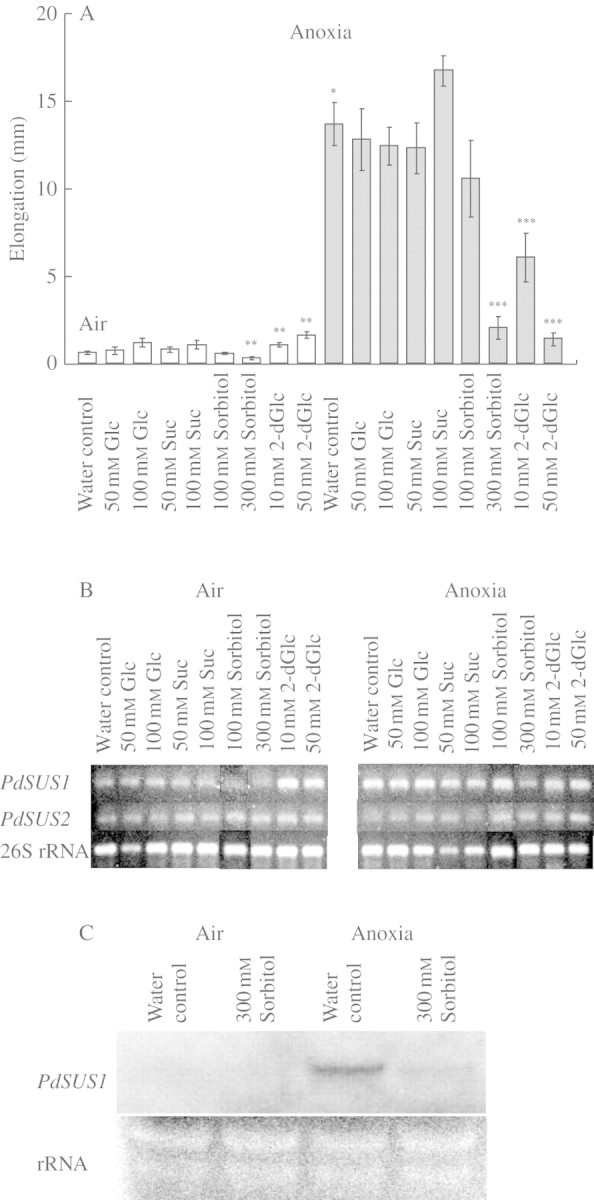
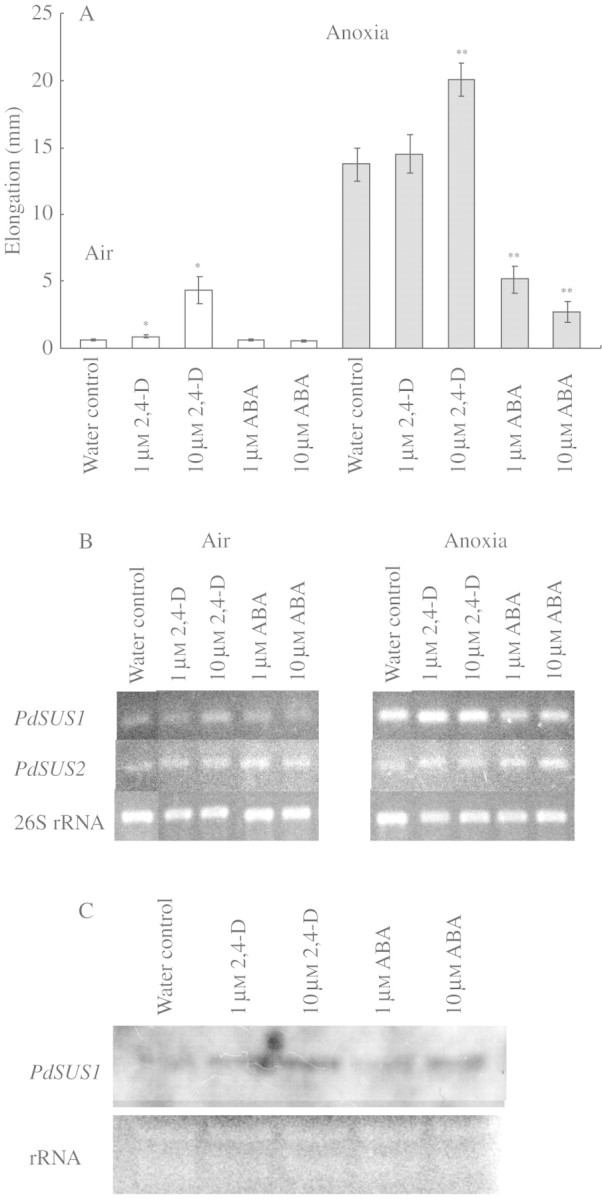
Key results: cDNAs of two SuSy genes (PdSUS1 and PdSUS2) were cloned from pondweed turions. The levels of PdSUS1 transcripts increased under anoxia but did not with sugar treatments. Anoxia-stimulated elongation of turions was further enhanced by 2,4-dichlorophenoxyacetic acid (2,4-D) and suppressed by treatments with sorbitol, 2-deoxyglucose (2-dGlc) and abscisic acid (ABA). The levels of PdSUS1 transcripts were increased by 2,4-D and decreased by sorbitol under anoxia. The levels of PdSUS2 transcripts were not significantly affected by anoxia and any other treatments. SuSy activity of turions under anoxia was enhanced in the soluble fraction, but not in the microsomal fraction.
Conclusions: Up-regulation of PdSUS1 transcription under anoxia may not be attributed to sugar starvation under anoxia. A positive correlation between stem elongation and the level of PdSUS1 transcripts was observed in turions treated with anoxic conditions, 2,4-D and sorbitol. The increase in SuSy activity in the cytosol may contribute to sugar metabolism and sustain stem elongation under anoxia.
Figures

Similar articles
-
Anoxia-enhanced expression of genes isolated by suppression subtractive hybridization from pondweed (Potamogeton distinctus A. Benn.) turions.Planta. 2007 Sep;226(4):1041-52. doi: 10.1007/s00425-007-0537-8. Epub 2007 May 15. Planta. 2007. PMID: 17503072
-
Involvement of plasma membrane H+-ATPase in anoxic elongation of stems in pondweed (Potamogeton distinctus) turions.New Phytol. 2011 Apr;190(2):421-30. doi: 10.1111/j.1469-8137.2010.03605.x. Epub 2011 Jan 14. New Phytol. 2011. PMID: 21232059
-
Stimulation of glycolysis in anaerobic elongation of pondweed (Potamogeton distinctus) turions.J Exp Bot. 2002 Sep;53(376):1847-56. doi: 10.1093/jxb/erf036. J Exp Bot. 2002. PMID: 12177122
-
Regulation of sucrose metabolism in higher plants: localization and regulation of activity of key enzymes.Crit Rev Biochem Mol Biol. 2000;35(4):253-89. doi: 10.1080/10409230008984165. Crit Rev Biochem Mol Biol. 2000. PMID: 11005202 Review.
-
Overexpression of PsnSuSy1, 2 genes enhances secondary cell wall thickening, vegetative growth, and mechanical strength in transgenic tobacco.Plant Mol Biol. 2019 Jun;100(3):215-230. doi: 10.1007/s11103-019-00850-w. Epub 2019 May 4. Plant Mol Biol. 2019. Retraction in: Plant Mol Biol. 2023 May;112(1-2):105. doi: 10.1007/s11103-023-01353-5. PMID: 31053988 Retracted. Review.
Cited by
-
Cloning, characterization, and expression of xyloglucan endotransglucosylase/hydrolase and expansin genes associated with petal growth and development during carnation flower opening.J Exp Bot. 2011 Jan;62(2):815-23. doi: 10.1093/jxb/erq319. Epub 2010 Oct 19. J Exp Bot. 2011. PMID: 20959626 Free PMC article.
-
Anoxia-enhanced expression of genes isolated by suppression subtractive hybridization from pondweed (Potamogeton distinctus A. Benn.) turions.Planta. 2007 Sep;226(4):1041-52. doi: 10.1007/s00425-007-0537-8. Epub 2007 May 15. Planta. 2007. PMID: 17503072
-
The sucrose synthase gene family in blueberry (Vaccinium darrowii): functional insights into the role of VdSUS4 in salt stress tolerance.Front Plant Sci. 2025 Jun 2;16:1581182. doi: 10.3389/fpls.2025.1581182. eCollection 2025. Front Plant Sci. 2025. PMID: 40530277 Free PMC article.
-
Genome-wide analysis of sucrose synthase family in soybean and their expression in response to abiotic stress and seed development.PLoS One. 2022 Feb 25;17(2):e0264269. doi: 10.1371/journal.pone.0264269. eCollection 2022. PLoS One. 2022. Retraction in: PLoS One. 2022 Sep 14;17(9):e0274409. doi: 10.1371/journal.pone.0274409. PMID: 35213642 Free PMC article. Retracted.
-
Structure, expression profile and subcellular localisation of four different sucrose synthase genes from barley.Planta. 2011 Aug;234(2):391-403. doi: 10.1007/s00425-011-1408-x. Epub 2011 Apr 20. Planta. 2011. PMID: 21505865
References
-
- Albrecht G, Mustroph A. 2003. Localization of sucrose synthase in wheat roots: increased in situ activity of sucrose synthase correlates with cell wall thickening by cellulose deposition under hypoxia. Planta 217: 252–260. - PubMed
-
- Balk PA, de Boer AD. 1999. Rapid stalk elongation in tulip (Tulipa gesneriana L. cv. Apeldoorn) and the combined action of cold-induced invertase and the water-channel protein γTIP. Planta 209: 346–354. - PubMed
-
- Baud S, Vaultier MN, Rochat C. 2004. Structure and expression profile of the sucrose synthase multigene family in Arabidopsis Journal of Experimental Botany 55: 397–409. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
