

Nonauditory events of a behavioral procedure activate auditory cortex of highly trained monkeys
- PMID: 16033889
- PMCID: PMC6725347
- DOI: 10.1523/JNEUROSCI.1571-05.2005
Nonauditory events of a behavioral procedure activate auditory cortex of highly trained monkeys
Abstract
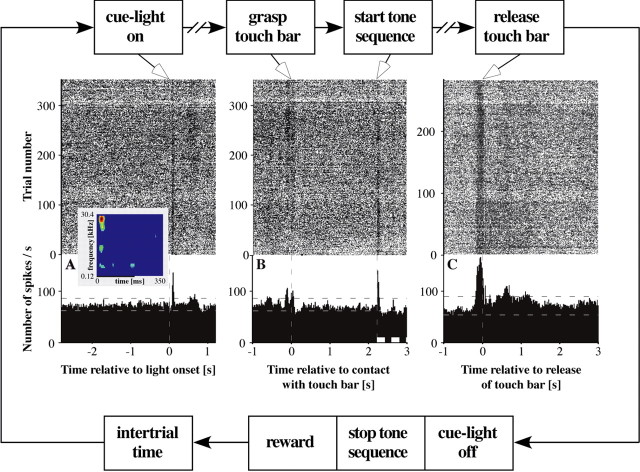
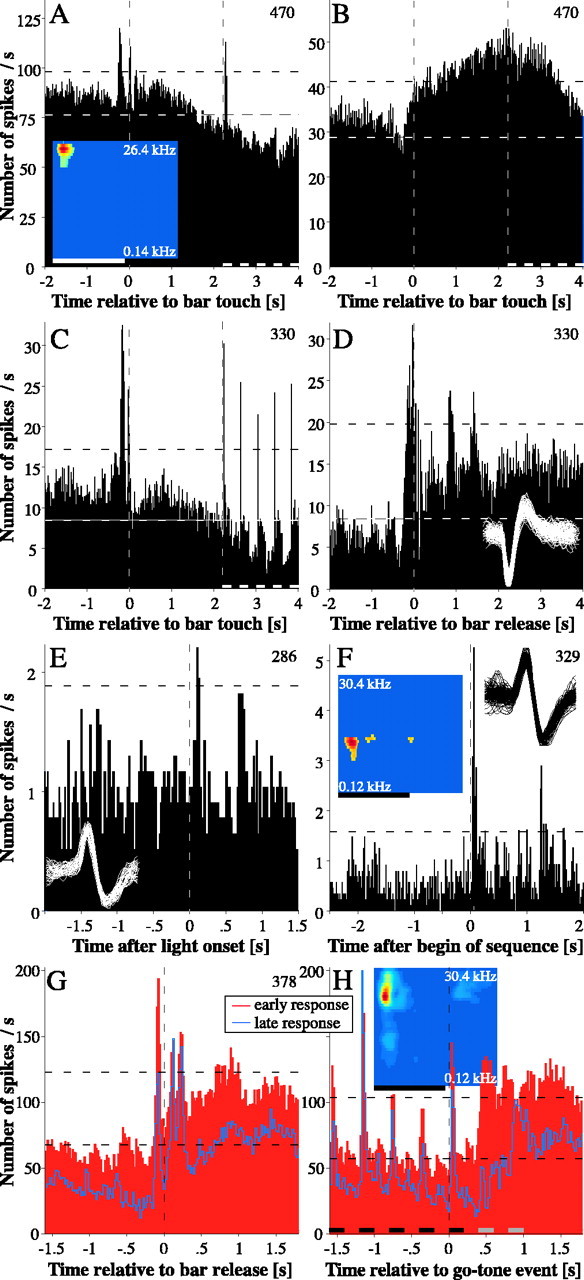
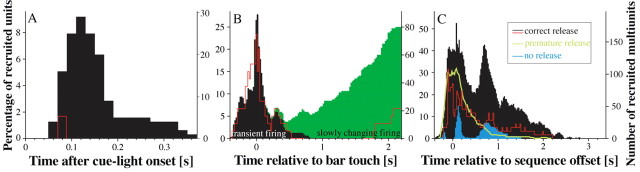
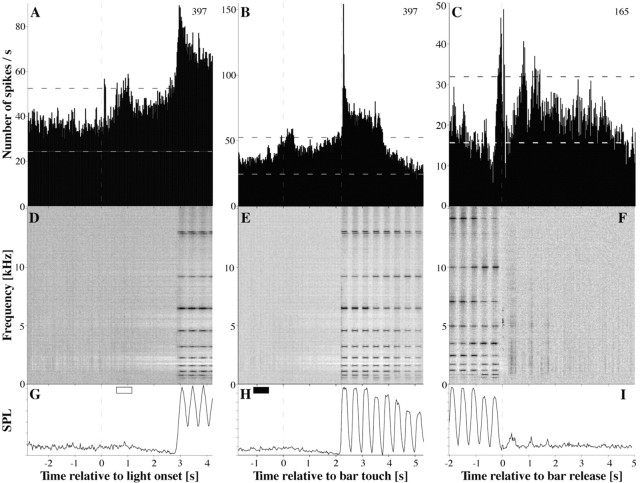
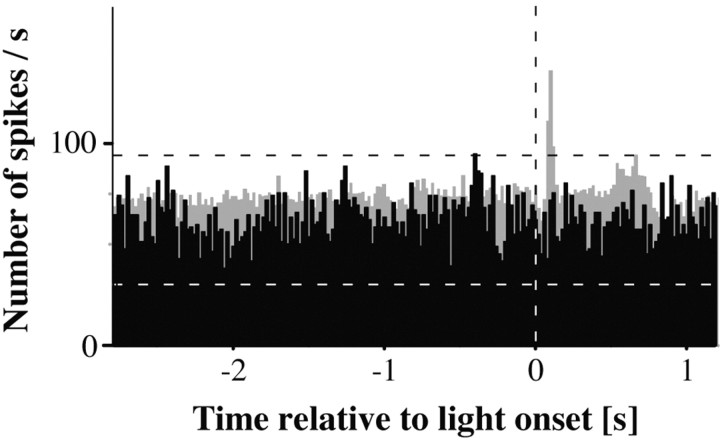
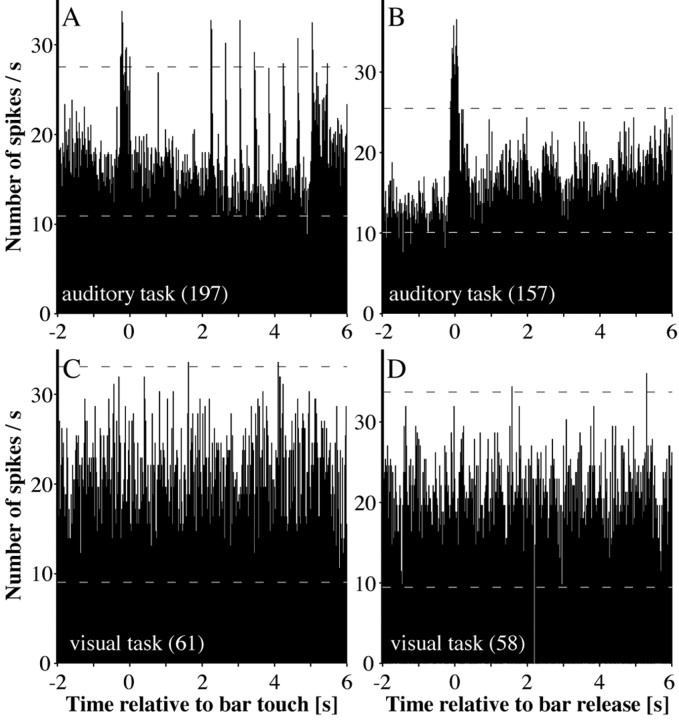
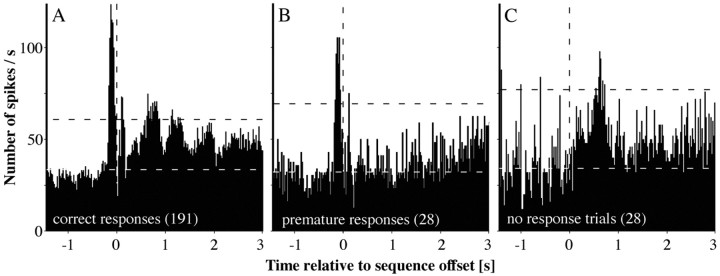
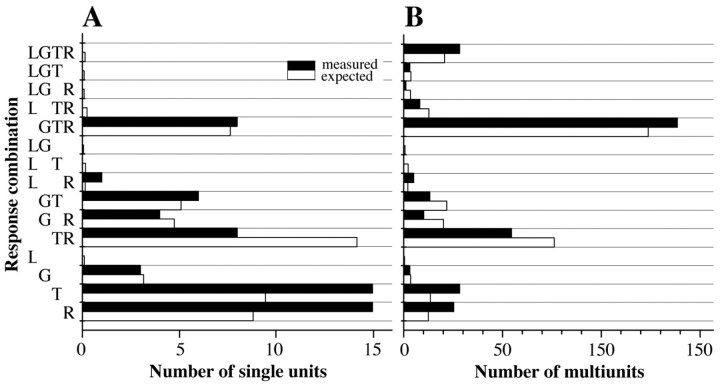
A central tenet in brain research is that early sensory cortex is modality specific, and, only in exceptional cases, such as deaf and blind subjects or professional musicians, is influenced by other modalities. Here we describe extensive cross-modal activation in the auditory cortex of two monkeys while they performed a demanding auditory categorization task: after a cue light was turned on, monkeys could initiate a tone sequence by touching a bar and then earn a reward by releasing the bar on occurrence of a falling frequency contour in the sequence. In their primary auditory cortex and posterior belt areas, we found many acoustically responsive neurons whose firing was synchronized to the cue light or to the touch or release of the bar. Of 315 multiunits, 45 exhibited cue light-related firing, 194 exhibited firing that was related to bar touch, and 268 exhibited firing that was related to bar release. Among 60 single units, we found one neuron with cue light-related firing, 21 with bar touch-related firing, and 36 with release-related firing. This firing disappeared at individual sites when the monkeys performed a visual detection task. Our findings corroborate and extend recent findings on cross-modal activation in the auditory cortex and suggests that the auditory cortex can be activated by visual and somatosensory stimulation and by movements. We speculate that the multimodal corepresentation in the auditory cortex has arisen from the intensive practice of the subjects with the behavioral procedure and that it facilitates the performance of audiomotor tasks in proficient subjects.
Figures
References
-
- Bangert M, Haeusler U, Altenmuller E (2001) On practice: how the brain connects piano keys and piano sounds. Ann NY Acad Sci 930: 425-428. - PubMed
-
- Beaton R, Miller JM (1975) Single cell activity in the auditory cortex of the unanesthetized, behaving monkey: correlation with stimulus controlled behavior. Brain Res 100: 543-562. - PubMed
-
- Benson DA, Hienz RD (1978) Single-unit activity in the auditory cortex of monkeys selectively attending left vs. right ear stimuli. Brain Res 159: 307-320. - PubMed
-
- Bronchti G, Heil P, Sadka R, Hess A, Scheich H, Wollberg Z (2002) Auditory activation of “visual” cortical areas in the blind mole rat (Spalax ehren-bergi). Eur J Neurosci 16: 311-329. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources