

Histone deacetylase inhibitors induce reactivation of herpes simplex virus type 1 in a latency-associated transcript-independent manner in neuronal cells
- PMID: 16036811
- PMCID: PMC1361429
- DOI: 10.1080/13550280590952817
Histone deacetylase inhibitors induce reactivation of herpes simplex virus type 1 in a latency-associated transcript-independent manner in neuronal cells
Abstract
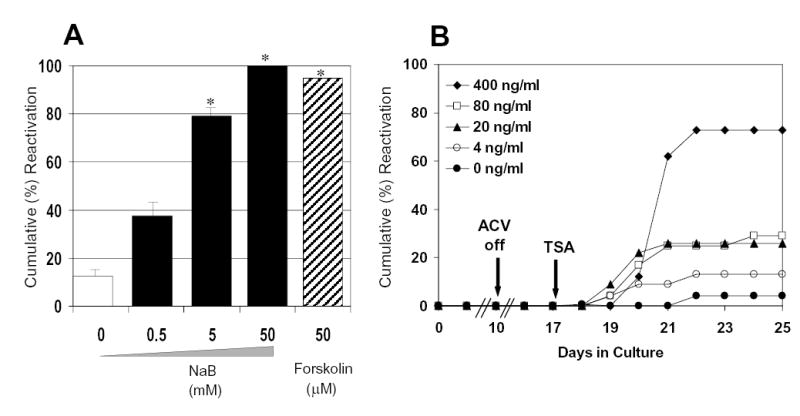
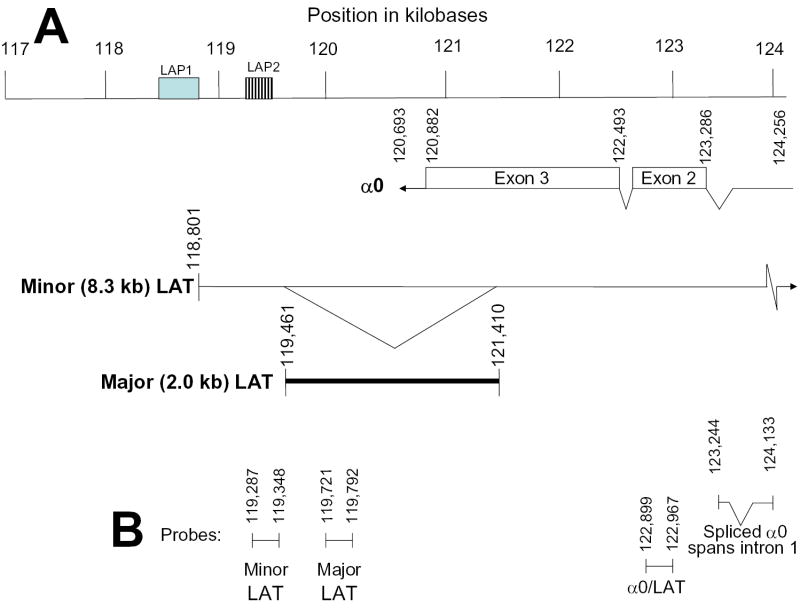
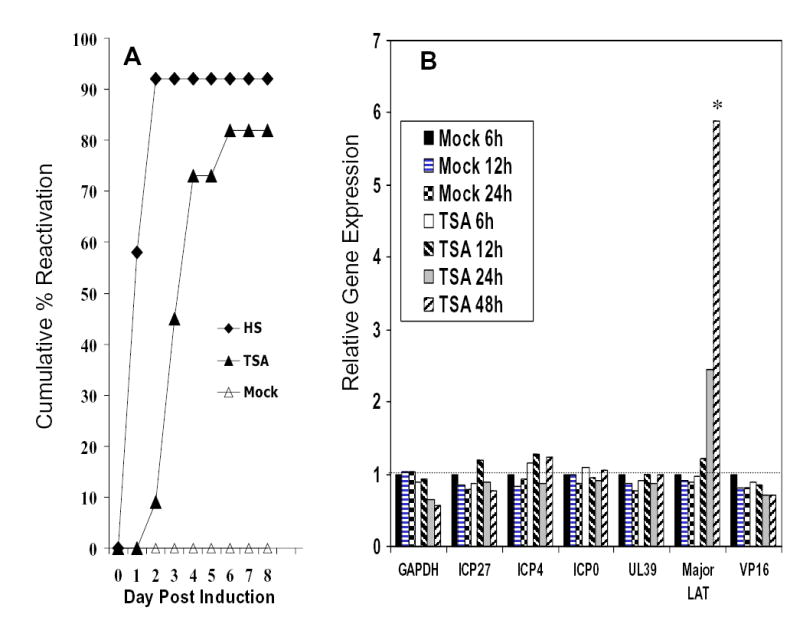
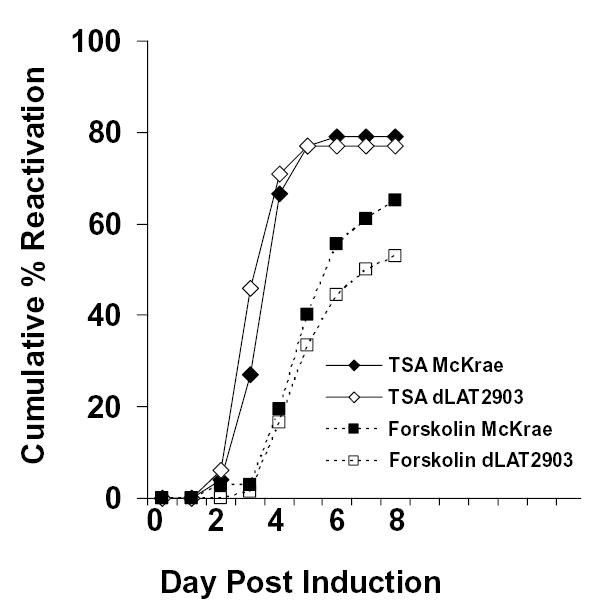
Histone acetylation is implicated in the regulation of herpes simplex virus type 1 (HSV-1) latency. However, the role of histone acetylation in HSV-1 reactivation is less clear. In this study, the well-established model system, quiescently infected, neuronally differentiated PC12 (QIF-PC12) cells, was used to address the participation of histone acetylation in HSV-1 reactivation. In this model, sodium butyrate and trichostatin A (TSA), two histone deacetylase inhibitors, stimulated production of infectious HSV-1 progeny from a quiescent state. To identify viral genes responsive to TSA, the authors analyzed representative alpha, beta, and gamma viral genes using quantitative real-time polymerase chain reaction. Only the latency-associated transcript (LAT) accumulated in response to TSA treatment, under culture conditions that restricted virus replication and spread. This led the authors to evaluate the importance of LAT expression on TSA-induced reactivation. In QIF-PC12 cells, the LAT deletion mutant virus dLAT2903 reactivated equivalently with its wild-type parental strain (McKrae) after TSA treatment, as well as forskolin and heat stress treatment. Both viruses also reactivated equivalently from latently infected trigeminal ganglia explants from rabbits. In contrast, there was a marked reduction in the recovery of dLAT2903, as compared to wild-type virus, from the eyes of latently infected rabbits following epinephrine iontophoresis. These combined in vitro, ex vivo, and in vivo data suggest that LAT is not required for reactivation from latently infected neuronal cells per se, but may enhance processes that allow for the arrival of virus at, or close to, the site of original inoculation (i.e., recrudescence).
Figures
References
-
- Bloom DC, Stevens JG, Hill JM, Tran RK. Mutagenesis of a cAMP response element within the latency-associated transcript promoter of HSV-1 reduces adrenergic reactivation. Virology. 1997;236:202–207. - PubMed
-
- Boutillier AL, Trinh E, Loeffler JP. Constitutive repression of E2F1 transcriptional activity through HDAC proteins is essential for neuronal survival. Ann NY Acad Sci. 2002;973:438–442. - PubMed
-
- Boutillier AL, Trinh E, Loeffler JP. Selective E2F-dependent gene transcription is controlled by histone deacetylase activity during neuronal apoptosis. J Neurochem. 2003;84:814–828. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
