

An RNA-dependent RNA polymerase prevents meristem invasion by potato virus X and is required for the activity but not the production of a systemic silencing signal
- PMID: 16040651
- PMCID: PMC1183376
- DOI: 10.1104/pp.105.063537
An RNA-dependent RNA polymerase prevents meristem invasion by potato virus X and is required for the activity but not the production of a systemic silencing signal
Abstract
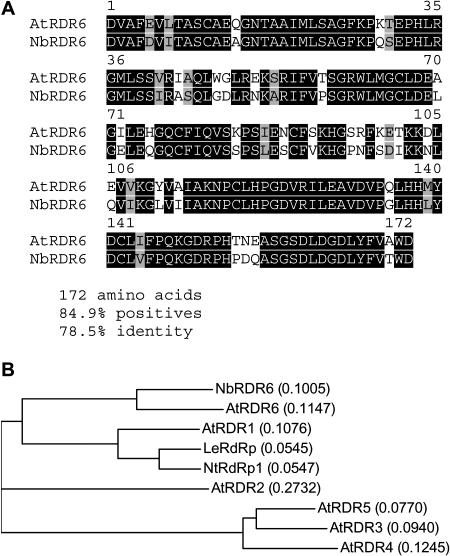
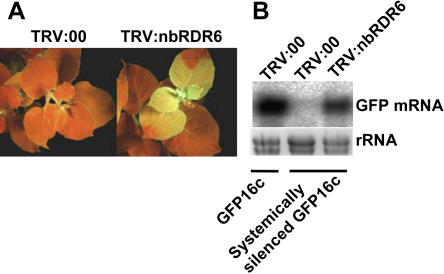
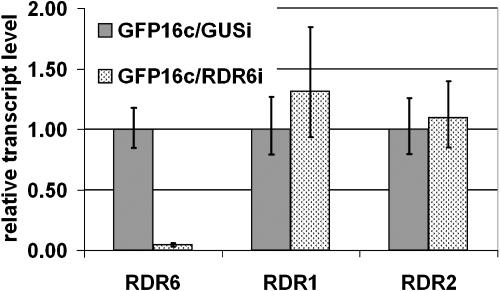
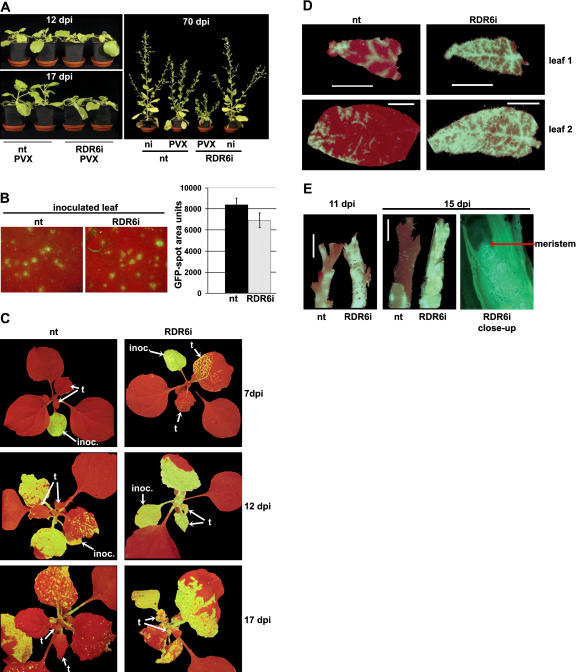
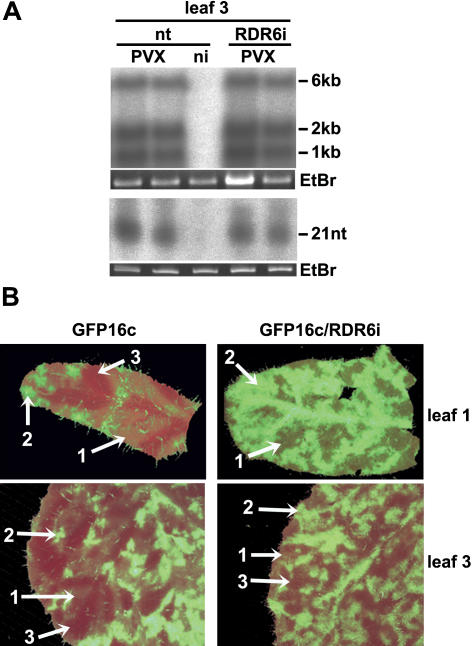
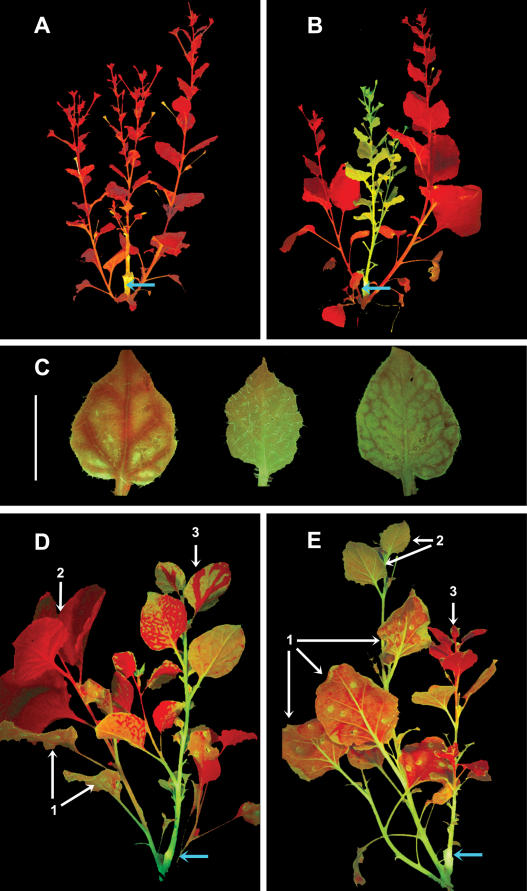
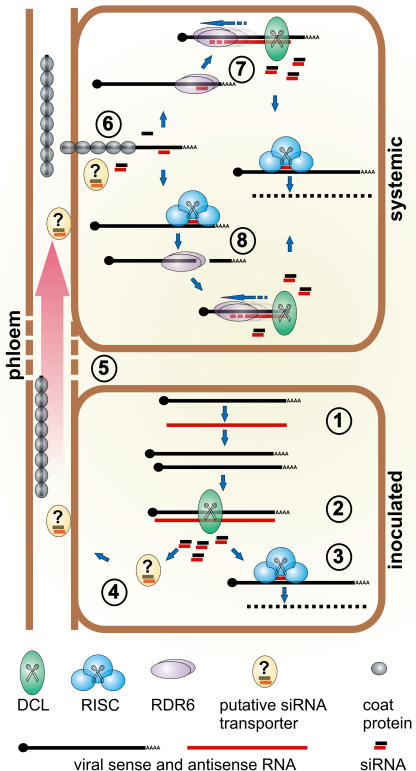
One of the functions of RNA silencing in plants is antiviral defense. A hallmark of RNA silencing is spreading of the silenced state through the plant. Little is known about the nature of the systemic silencing signal and the proteins required for its production, transport, and reception in plant tissues. Here, we show that the RNA-dependent RNA polymerase RDR6 in Nicotiana benthamiana is involved in defense against potato virus X at the level of systemic spreading and in exclusion of the virus from the apical growing point. It has no effect on primary replication and cell-to-cell movement of the virus and does not contribute significantly to the formation of virus-derived small interfering (si) RNA in a fully established potato virus X infection. In grafting experiments, the RDR6 homolog was required for the ability of a cell to respond to, but not to produce or translocate, the systemic silencing signal. Taking these findings together, we suggest a model of virus defense in which RDR6 uses incoming silencing signal to generate double-stranded RNA precursors of secondary siRNA. According to this idea, the secondary siRNAs mediate RNA silencing as an immediate response that slows down the systemic spreading of the virus into the growing point and newly emerging leaves.
Figures

References
-
- Astier-Manifacier S, Cornuet P (1974) RNA-dependent RNA polymerase in Chinese cabbage. Biochim Biophys Acta 232: 484–493 - PubMed
-
- Baulcombe D (2004) RNA silencing in plants. Nature 431: 356–363 - PubMed
-
- Baulcombe DC, Chapman SN, Santa Cruz S (1995) Jellyfish green fluorescent protein as a reporter for virus infections. Plant J 7: 1045–1053 - PubMed
-
- Beclin C, Boutet S, Waterhouse P, Vaucheret H (2002) A branched pathway for transgene-induced RNA silencing in plants. Curr Biol 12: 684–688 - PubMed
-
- Bernstein E, Caudy AA, Hammond SM, Hannon GJ (2001) Role for a bidentate ribonuclease in the initiation step of RNA interference. Nature 409: 363–366 - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
